�����ڡ����ڷ��ڣ������ڷ��ڣ�ϵͳ������ϵͳ�ĵ���
 |
| ���ڹ�����ѧϰ����ģʽ/ |
 |
| ����ϵͳ�ڵĹ�����ѧϰģʽ/ |
�������ڷ���ѧ��һ��Ҫ���������ڷ���ϵͳ(�����ڷ���)�����߹��ܵĵ��ء������Ͻ������е��ڷ��ڹ��ܾ�����ϵͳ��ֱ�ӻ���֧�䣬�����ڷ���ϵͳ�������ڷ��ڱ�ʾ�����ڷ��ڶ�����ϵͳ��Ӱ�����ɼ��ء����ġ����ʵ�������ʵ�֣�������һЩ���͵��������̻�ʵ������У���Ӧ����������������������Ӧ��IJ����ȡ�
һ�����ڷ��ڶ����߹��ܵ��ص�����ѧ����
�ܰ���֯���ܰ����پ�����ȥ���Ǿ�����֪����ʵ����Щ����ά����Ѫ�ܴ�����Ĥ�������ܰ���֯��������Ϊ���л�������ά�������������������֧������������ܰ����ٵ������ڶ���������ά��
1.���� �����е�������ά��Ƕ���������������裬֧�������Ѫ�ܼ�ʵ�ʣ���ϸ����ϵ���С���Щ��ά���������������ά��������P����(substance P,SP)��������ά��
2.���� ��Ϊ�����������ٺ��ڷ������٣����ٿɽ�������������������֧�䣬���н�������ά��Դ�ھ��ضν�������������������ά��Դ����������������֯��ѧ�����ٷ�֤ʵ��֧�����ٵ���������ά������������ˡ�ӱ�ˡ��������˵Ⱥ��ţ���������״ϵͳ�Ĵ���嶯���������乹�ɶ�ͻ����ϵ����Щ������ǰ��Ԫ���и��Է���(leucine-enkephalin,L-ENK)����������(SS)���ڷ������ڣ���������ά���ֲ������٣������γ���������Щ����ά������ķ����������˥�˶��仯�����к���������ά��Ƥ���гɴԷֲ�������ĩ�ҽ������ʣ������ϸ������ڡ�Ѫ�ܻ��Գ���(vasoactive intestinal polypeptide,VIP)�������ػ��������(calcitonin-gene-related polypeptide,CGRP)������Y(neuropeptide Y,NPY)���Ե��ֲ��ڴ�������Ѫ����Χ��СҶ������֯��Ƥ�ʺ����ʵ�ʵ���ڡ���Ĥ����SP��CGRP������ά��ȥ����������(norepinephrine,NE)������ά�ֲ������з�֧����������ʵ��ϸ���䣬��ʴ�ϸ��������ϸ����ϵ���С��ѷ���NPY�����NE����ĩ���С���Щ��ʵ˵�����ٵĽṹ�ͻ��ܿ��ܽ��к��������Ӱ�졣һ����Ϊ�������˷ܿɼ��������ܣ������������˷��������෴��
3.Ƣ ���Ը�ǻ�ڵĽ������γ�Ƣ����Ƣ����Ƣ�������鶯����Ƣ������Ƣʵ�ʰ����С�����������ܰ�ϸ����ϵ���У������ڰ���ͺ���Ľ��紦������Ƣ��������붯�������֧����NPY���������Է���(methionine-enkephalin,M-ENK)��������(cholecystokinin,CCK)����ѹ��(neurotensin,NT)��������ά�ֲ�������������ʵ�ʡ�èƢ��Ѫ�����ຬ��NPY��SP��VIP������������ά���ѷ���NPY��NE���档������ά��Ҫ����Ƣ�౻Ĥ��С����λ��Ѫ�ܡ������Ұ����ǻ�ø(tyrosine hydroxylase,TH)������ĩ�ҿ���Ƣ�ܰ�ϸ���γ�ͻ����ϵ��
4.�ܰͽ� ���ܰͽ�İ�Ĥ�¼���Ĥ�ڣ��ɼ���������(acetylcholine,ACh)������ά����NE�������ܰͽ�ʵ���У����ܱ�Ѫ�ܳɴԷֲ���������ʵ�������롣���ܰͽ���Ų�����Ĥ�¡�Ƥ�������ʽ��缰���ʺ�Ƥ������SP��VIP��NPY��CGRP����������ά�ķֲ���
5.�ܰ� ���к���������ά֧���ܰܣ������������ص㡣���⣬����ճĤ�²���ܰ�С���Peyer�Ͻ���ճĤ����������أ�����SP��������ά��֧�䡣SP��������ά�������ڳ���ë�������ӹ���Χ��������ACh������ά�ֲ���
��������ʵ˵����������֯�������ܵ�����������������������ά��֧�䣬����̬�����ֳ���ϵͳ������ϵͳ��ֱ��Ӱ�죬������֧�����ͻ����ʽ�ͷǵ���ͻ�����ַ�ʽ���������Դ������͡�����Դ�����������Ӧ��Ϊ����άĩ�ҵ��Է������������������������ַ�ʽ����Ϊ�����в�֪��������ά���ܰ���֯�����ٵ�Ӱ�������漰���¼����棺��Ѫ�����أ����ܰ�ϸ���ķֻ������������졢���к���ѭ������ϸ�����ӻ������������ӵ����ɺͷ��ڣ�������Ӧ���ǿ����ά�ֵ�ʱ��ȡ�
����ҩ��ѧ���������������������������Ӱ�����������������������ѧ�ȼ�������������ϸ��Ĥ�ϻ���ڷ����ڶ༤�ء����ĺ����ʵ����������塣
1.������������
(1)�����������壺��������������ɷ�Ϊ���ͦ����ͣ������ٷ�Ϊ��1����2�ͦ�1����2�͡���֪���ٺ�Ƣ����������������ά��֧�䣬������������ϸ����Ƣϸ��Ĥ���Ц�����ķֲ���С������ϸ������������̥�ڵ�����(Kd=2.2nM)���ڳ�����(Kd=8.0nM)���æ����弤��������������ش̼���̥������ϸ��cAMP����Ҳ��Ϊ���ԡ�С���ܰ�ϸ���Ϧ������KdΪ1nM�����λ����Ϊ500��/ϸ�������б�������125I��ǵ��������������������������С���ܰ�ϸ��������Ϊ��2���ͣ���Kd=0.9nM�������ܶ�Ϊ3000��/ϸ������24�����ϵС����е��������������T��Bϸ���Ϧ�2���嶯��ѧ����������ͬ������Ա�Ӱ�졣
����ǻ�ʴ�ϸ��Ĥ���Ц��ܱ��ֲ���KdΪ1.58��0.56nM���������Ϊ4000��14000��/ϸ����ҩ��ѧ������������Щ��������83.5%Ϊ��2����16.5%Ϊ��1���ͣ�����IgE���������Ag�̼��ʴ�ϸ�������ı����������ԡ���ֹ�ķʴ�ϸ���Ԧ����弤�����̼�15�룬����cAMP�������Ե����࣬����Ӱ��IgE�鵼����֯�����ͷš���������Ƣʵ��ϸ���Ц�2�ͦ�2����Ĺ��档
��������Ѫ�ܰ�ϸ���Ħ������ܶ�Ϊ3500����/ϸ������KdΪ0.35��0.18nM��
������Ѫ�ܰ�ϸ�������κ˰�ϸ��������ϸ��������ϸ����ѪС���Ͼ����������������塣�����˺����������ܰ�ϸ���Ϧ������ܶ����죬�������������䣬���æ����弤�����ɽ��ͦ������ܶȣ����µ�����������Ը������ÿ�����terbutaline 6mg��7����ܰ�ϸ���Ϧ�2������Ŀ����42%��T��B�ܰ�ϸ����������������Բ��졣ѪС�����Ц��������������壬�̼�������ɴٽ�ѪС��ۼ����˶��κ˰�ϸ����M�����Ц�2����Ĵ��ڡ������½����������Ը�Ⱦ��������Ѫ�е�����ϸ���Ħ�2�����ܶ���Ѫ���п�����IL-2�����ˮƽ�йء�
(2)��Ͱ����壺С��ʹ����ܰ�ϸ��Ĥ�ж�Ͱ����壬��Kd�ֱ�Ϊ4.8��0.2nM��1.9nM��С��B�ܰ�ϸ���ϵ������ܶ�Ϊ60000��/ϸ�����Ҷ��ֿ���Ͱ�ҩ��������Ʒ�����������Ͱ�����Ľ�ϡ�
(3)ACh���壺��ͬλ�ر�ǵ�M����ɼ���С��Ƣ�ܰ�ϸ���ͷ��ܰ�ϸ���ϣ�ǰ���ǵ�KdΪ1nM���ܶ�Ϊ200��/ϸ���������ܶ�Ϊ400��/ϸ����С�����ٺͼ�������ϸ���ĵ���������ΪN�ͣ�������ؽ�����������أ���ʾ���ٵIJ���ϸ��������Դ�����ջ������߲㡣�����ϸ��Ĥ�ϵ�ACh��������ϸ���ļ�����⣬Jurkatϸ�������M3�͵�ACh���壬M3���弤���ϸ����Ca2+Ũ����������һЧӦϵPLC��IP3���鵼�ġ�Jurkatϸ��Ĥ��M3�����KdΪ14.1nM�������Ϊ45370λ��/ϸ�������Ʊ�ϸ��ĤҲ������һ��ϻ��ԡ�
(4)5��ɫ(5-hydroxytryptamie,5-HT):5-HT�������ʣ���������ϸ����ʴ�ϸ�����ȼ�����ϸ���Ļ��Է�����ѷ��ּ������Tϸ��Ĥ����5-Ht 1a����ķֲ���������ļ�������Ӱ���cAMP��Ũ�ȡ���T�ܰ�ϸ��ϵJurkatҲ����5-HT1a���壬�����ÿ���IP3��Ca2+�鵼��
(5)��֯��(histamine)���壺��Tϸ����KdΪ0.4nM����ĿΪ35000/ϸ������֯�����塣CD8+Tϸ���ϵ��鰷����ΪH2�ͣ����ܶ�ԼΪ6000-7000/ϸ�������ܰ�ϸ�����غ�H2�������������涡�ĵ��ڡ�Bϸ���ϵ���֯����������H2�;��жࡣ
2.��̴���������
��������ѧ�о�����������������̴����صİ������幹��һ�ܱ������壬������Ƥ�ʼ�������(glucocorticoid ,receptor,GR)���ۼ�������(androgenreceptor,AR)���м�������(progesterone receptor,PR)����Ƥ�ʼ�������(mineralocorticoid,receptor,MR)����״�ټ�������(thyroidhormone receptor,TP),�ӻ�������(retinoicacid receptor,RAR)��ά����D����(vitaminD erceptor,VDR)������GR��MR��AR��PR����ʶ��ͽ��DNA������һ�λ������У���֮Ϊ��Ƥ�ʼ��ط�Ӧԭ��(glucocorticoid response element,GRE),������ΪAGAACAnnnTGTTCT��GRE�鵼��Ӱ��л����ת¼��
(1)GR��GR�㷺�ֲ��ڶ����ܰ���֯�����٣�����������ϸ���İ��������ڡ�GR������ϸ���ı����������ص㣺���нϴ���Ա���죬�������ϸ����GRŨ�ȵ������ڿ�ԭ�̼���PHA�����ϵ�GRŨ�ȣ�������������ʹ���µ�����Ca2+�����GRʧ�����ʵĵ��ء���IL-2��IL-4����Ӧ�ÿɽ���������Ѫ������ϸ��GR���������������ʹ�����ӣ������ÿɱ�IFN-�����ϡ���LPS�ɴٽ�GR��С��M�յı��
GR��800�ఱ������ɣ������пɷ�Ϊ������������DNA���������80������л����ɣ����IJ���Ϊ����пָ�ṹ���ɽ��DNA˫�����Ĵ�(major groove)�ڣ���C��Ϊ��Ƥ�ʼ���(GC)���������Nĩ����Tau1��������GR��DNA��Ϻ��Է�ʽ����л����ת¼�����⣬��GC������ڽ���Ҳ��Taul��������GR�ں���תλ�����������ʱ��GR��300kDa�ĵ��������϶���������״̬���˵���������2���������90kDa���ݿ˵���(HSP90)��59kDa����������(immunophilin)�����������Ե�����ɣ�HSP90�����GR��Cĩ�ˣ�����GR���۵����Ͳ���ֹ�����������תλ����������Ҳ�ɽ��CsA��FK506��rapmycin���������Ƽ�����ʾGC������ЧӦ�������������Ƽ������û��ƿ�����ͬ��
(2)AR����֤ʵ�ڴ���С������ټ����ķ�������Ƥϸ������غͪ������ڡ�
(3)ER��PR���ƶ���(F2)��������ڴ���С��ţ���˵������У���KdԼΪ0.2nM�����ܶ�λ��������Ƥϸ������״ϸ���ڡ���������Ҳ��E2���塣PR����ڴ�������ϸ���ڡ�
3.���ļ����༤������
(1)ACTH���壺��֪С��Ƣϸ��Ĥ���и������͵���������ACTH���壬��Kd�ֱ�Ϊ0.1nM��4.8nM�������ܶ�Ϊ3000/ϸ����5000/ϸ����������Ѫ������ϸ��Ҳ�д��������壬Kdֵ�ֱ�Ϊ0.04M��3.4nM������ϸ���ϵ�ACTH������������Ƥ��ϸ��Ĥ��ACTH�������ʺͽṹ������ͬ���������Ϊ225kDa����4�����������ɣ��������ֱ���83��64��52��22kDa��
(2)GH���壺С��Сţ����ϸ������Tϸ��������Ѫ������ϸ������GH����ֲ������ٵ�GH�����ܶ�Ϊ10-20000/ϸ������IM-9�ܰ�ĸϸ��ϵϸ��Ĥ��GH�����KdΪ1.3nM���ܶ�Ϊ4000/ϸ����
(3)PRL���壺PRL��GH������ϸ�����ù㷺���������������Ӧ����������ϸ��Ĥ�ϡ�������ѪT��B�ܰ�ϸ��PRL����Kd=1.66nM���ܶ�Ϊ360/ϸ������E2���µĴ����ܰ���Nb2��ϸ����PRL����KdΪ75pM��ÿ��ϸ����12000�����λ�㡣�����֣�����ù��A�������ܰ���ϸ����125I-PRL�Ľ�ϡ�
(4)��Ƭ�����壺��Ƭ�����岻ͬ�����ͼ��ǰ�Ƭ�����������������ϸ��Ĥ�ϡ�������Ѫ�ܰ�ϸ����ѪС��ɽ��3H-����ͪ���ǰ�Ƭ��������Ҫ���-�ڷ���(��-endorphn,��-END)��C�����ϣ���������ܰ�ϸ����ֲ��Ѫ���صķ�Ӧ��
(5)SP���壺���÷��������������������Ӱ��FACS�ȼ�����֤ʵSP����������ֲ��㷺�������Ƣ���Bϸ���ı�Ե����С��Ƣ�༰����Peyer�Ͻ��е�T��Bϸ��������ǻ����ϸ����(��10-2)��
��10-2 SP����������ϸ���ϵİ䲼������
�����ϸ����Դ
�������� ��
�������볣��(Kd)
�����ܶ� (��/ϸ��)
��������ʶ��λ
������ѪT�ܰ�ϸ��
3H-SP
1.85��0.70*10-7
7035��2850
C-ĩ��
��BϵIM-9�ܰ�ĸϸ��
3H-SP
0.65��0.19*10-9
22641��6143
C-ĩ��
��BϵIM-9�ܰ�ĸϸ��
ϸ��Ĥ���
125I-SP
0.87��0.80*10-9
21��3*10-15M/mg����
C-ĩ��
��Ĥ������
125I-SP
0.75��0.33*10-9
3.7��1.5*10-15M/mg����
C-ĩ��
С���ܰ�ϸ��(���)
0.68��0.01*10-9
N��Cĩ��
Ƣ�ܰ�ϸ��
125I-SP
195(T),190(B)
Peyer�Ͻ��ܰ�ϸ��
125I-SP
647(T),975(B)
����Ƣ���Ե��
125I-SP
2.4*10-9
ע�����ټ��������У�SP����ΪNK-1�ͣ�ϵG������������Ա֮һ��
(6)������CD4+Tϸ��Ĥ�����������ỷ��ø��ż����VIP���塣����������ֲ���������Bϸ��Ĥ�ϣ���ʾ�����ز����ܵ��ڹ����������̣�����Ӱ�����߹��ܡ�ͬ����LHRH����������Ƥ�ʼ����ͷż���(corticotropin-releasing hormone,CRH)��CGRP��ANP���ķ�������(atrial natriuretic polypeptide,ANP)��Ѫ�ܽ����آ�(angiotensin ��A��)��VIP��SS������Ҳ����������֯������ϸ���ϡ�
4.�ʺ������� Ŀǰ���֣��ڼ���Ѽ�ȼ��ݼ����ȵ����١�Ƣ�ࡢ�ܰͽ�ͷ����ҵȲ�λ�������Խ���ʺ��ء��ʺ��ؾ���֬���ԣ��ɴ���Ĥ�������ڰ������������塣
1.POMC���� ǰ����Ƥ��(proopiomelanocortin,POMC)Ϊ��������Ƥ�ʼ���(adrenocorticotropin,ACTH)��ǰ����ӣ�Ҳ�Ǧ�-LPH(��֬����)����-MSH(����ϸ���̼���)����-END��ǰ����������Ѫ�ܰ�ϸ����Ƣϸ���ڲ�����Ⱦ��LPS�����£��ɷ���ACTH�ͦ�-END���봹����ڵ�ACTH�ͦ�-END�ṹһ�¡�С��Ƣ���в���M�ռ�����ijЩ�ܰ�ϸ�����ȶ�������Щ���أ�POMc mRNA�ɱ�����M�ռ��ܰ�ϸ���С���PHA�̼��£�������Ѫϸ����POMc mRNA������ǿ��LPS�̼�С��Ƣϸ������ʹ�������POMC mRNA���ࡣС���ϸ���Բ�ͬ�Ĵ̼���Ӧ���죬����CRF���³Dz�����������Һ�У����ϸ�����ڵ�POMC������ACTH(1-39)�ͦ�-END(1-31)Ϊ������LPS�����ϸ����Ҫ����ACTH(1-25)�ͦ�-END(1-16��1-17)�����˽���-END��Ϊϸ�����ӡ�
2.TSH ��֤��������ѪBϸ����SEA�̼��¿ɷ���TSH��Molt-4����Tϸ����Ѫ��ϸ���꣬���ȶ�����TSH���仯ѧ�ṹ��������TSH��ͬ��
3.GH��PRL Con A������ܰ�ϸ����GH��PRL��mRNAˮƽ��T��Bϸ��Ҳ���ȶ�����GH���������Ϊ������߷�����������Ѫ���ܰ���֯��IM-9�ܰ�ĸϸ��ϵ��Jurkatϸ�������п���GH��PRL�����ת¼����Pit-1/GHF1 ,��Щϸ���ຬ��GH��PRL�ķ��Ӽ�mRNA����ˣ�GH�����Է��ڷ�ʽ�����ܰ�ϸ������Ѫϸ������ֳ�ͷֻ���GH����Щϸ���еĺϳɼ����ڿ�����Ҫ�ֲܾ��������Ӽ���̴����صĵ��ء�
4.SP ����������ϸ���ɺϳ�SP��Ѫ�������µ�С�������ѿ����������ϸ��Ҳ�ܺϳ�SP��
5.VIP��SS ��ѪС�塢������ϸ�����ʴ�ϸ��������ϸ���о�������VIP��SS�������������ʷֲ���
6.LHRH ����Ƣ�༰���ٵ��ܰ�ϸ���к�LHRH����mRNA������ϸ����mRNA���������������д��ڵ�LHRh mRNA�ĵ�������ͬ��Con A�ɴ̼�LHRH��T�ܰ�ϸ���ĺϳɣ�PHA�����������á��½����֣�������Ѫ��CD4+��CD8+Tϸ���ɱ���LHRH����mRNA������ˮƽ��ϸ��״̬��Ӱ�졣
7.CRH CRH��CRH mRNA�����ڴ������ٺ�Ƣ���У�������������CRH����mRNA�ṹ��������ͬ�����ٺ�Ƣ���е�CRH���������������Ϊ1.7:1:117��CRh mRNA��Ҫ���������ټ�Ƣ���е��ܰ�ϸ���ڡ���Щϸ����CRH���ڲ���IL-1��Ӱ�죬��֬����ø���Ƽ���ɼ�ǿCRH�ķ��ڣ���Щ������������CRH�ķ��ڵ��ڲ�ͬ��������Ѫ�ܰ�ϸ����������ϸ��Ҳ��CRH���Ӽ���mRNA�ı���ڴ�����֢��֯���ʪ�Թؽ��һ���ϸ����������ϸ��Ҳ��CRH���Ӽ���mRNA�ı���ڴ�����֢��֯���ʪ�Թؽ��һ�Ĥ��֯�������CRH�ķֲ���������ʵ��ʾCRH�����Է��ڻ��Է��ڵķ�ʽ�������ߵ��ء�
8.���� ������Ƥϸ�����ɺϳɾ�����Ѫ�ܼ�ѹ��(arginine vasporessia, AVP)���߲���(oxytocin, OT)��GHRH�ڴ����ϸ���еĺϳ���õ�֤ʵ��
�������ء����ļ����ʶ����߹��ܵ�Ӱ��
���ء����ļ����ʵ����ڷ�����Ϣ���ӿɽ辭���ڷ��ڡ��Է��ں��Է���;����Ӱ����������Ӧ�𣬲�����ijЩ���߲������̡�
1.��Ƥ�ʼ���(GC) GC�����߹��ܵ�Ӱ�켫Ϊ�㷺��Selye����1936�����ȹ۲�һ������Ƥ����ȡ��ɵ��´�������ή�������֤��GC��ͨ������;��Ӱ������ϵͳ���Ҵ�ЧӦ���ڽϴ���������죬��С��������ý�Ϊ���У������������Բ����У���Ҫ�������Ϊ���ߵ��ܰ�ϸ�����ױ�GC���÷����ܡ�
(1)GCӰ����̥������ϵͳ�ķ�������С����̥������GC�����£����ܰ�ϸ������Thy1.2��ԭ���ӣ�����Ũ�ȵ�GC��ɱ��С�ܰ�ϸ�������ٵ���Ƥϸ����GC��Ϊ���С�GC��������ή���Ļ����漰ϸ�����������������(apoptosis)����������Ҫ��������ø�IJ��룬��Ca2+��Mg2+�����������ø������DNA�Ķ��ѡ�GC���Խ������ټ��صķ���ˮƽ���������������ռ�ø(TK)�Ļ��ԡ������������ϸ���Ӵ�Gc 4Сʱ������RNA�������ʼӿ졣���⣬����ϸ����ͬ�����ζ�GC�ķ�Ӧ�������죬������̥�����У�����ǰ��ϸ����GC�������Խϳ���ϸ��Ϊ�ߡ�
(2)GCӰ���ܰ�ϸ�������ɺ�����Ѫ���ܣ�����ٹ����г���Bϸ����Ŀ�����������M�ռ���ϸ���ļ����γ��ʡ�
(3)GC�ı�ϸ����ѭ�������·ֲ���С�����GC��Ѫ�е�����ϸ������������ϸ�����١�GC�ɽ����ܰ�ϸ����Ѫ�н����ܰͽ�����������ٽ��ܰ�ϸ���չ�Ѫ����Ƥ���������ǻ��
(4)GC���ܰ��ܵĵ������ã���GC�ɽ���PHA�����Tϸ����ֳ��Ӧ��������뽵��IL-2R�����йء�GC���ܼ���Tϸ���������������ԣ�����Ƣ���е�Bϸ����LPS��PPD�ķ�Ӧ������Ig�ϳ�ϸ������Ŀ���ı�PWM�յ���PFC�γ��ʡ�����GC������M�յ�APC���������ƣ���IL-1���ڼ��١�GCҲ���Ƶ���ϸ��ת���M�գ�����Ƥ��Langerhansϸ���Ĺ��ܣ���������M�յ����ɼ�ϸ����ɱ���������۵���NKϸ���Ĺ��ܣ���С��Ƣϸ��������NKϸ�������ܵ�������(10-7-10-11M)�����ơ�����ҩ��������GC�����Ƣ����NKϸ����Ŀ����Ҳ���෴�ı�����
(5)GC�Էʴ�ϸ�����ܵ�Ӱ�죺GC����Ag�����P��ϸ���ѿ�����Ӧ��������֯�����ͷţ�������������ϸ����Ŀ��������������Ӧ��������ϸ�������������ɻ�����ܶԿ�ijЩϸ������ά����������ϸ���Ĵ�����á�
(6)GC��ϸ�����Ӳ�����������Ե�Ӱ�죺GC�ڷ�ʪ�Թؽ����߿�����IL-1�����IL-6����������LPS�յ���TNF-������������IL-����IL-1�¼�IL-6mRNA�IJ��ȶ��ԣ�����IL-2�ķ��ڣ���GM-CSF����Ӧ�ÿ����������ϸ����IL-1�Ľ�ϣ�����Ѫ��������Эͬ�յ�����ϸ������IL-1����GCͨ������IL-2R���źŴ��ݡ���������ϸ����IL-2�ķ�Ӧ�ԡ�GC��������RV38486��ģ��IL-1���ö���ǿTNT-��������������IL-6ЧӦ��GC���ɶԿ�IL-1�¶��ȵ��ط��ڵ�����ЧӦ��
(7)GC��MHC I�༰������ӱ�����������ã�MHC����Tϸ��ʶ��ճ����APC���ܣ�MHC������ͻ�����������ص�����ȱ�ݣ������������ɾ���Ѫ�������Լ���������֤����GC������С��Bϸ���;���ϸ��MHc �����I-A�µļ��GC�IJ�����������ЧӦ����֬Ƥ��(lipocoritin)�鵼�ģ�֬Ƥ����������6�ֵ����ʹ��ɵļ��壬��ṹ��ϸ���ǼܺͰ�����ص���(calpactin)���ơ���GC�����£�����ǻ��ϸ��������Ĥϸ����������Ѫ������ϸ������֧���ܷ��ݳ�ϴҺ��M�յȵ�֬Ƥ���������ӡ���ժ�������ٿɼ��ٴ�������֯��֬Ƥ��mRNA������ˮƽ��GC��ҪӰ��֬Ƥ��1��2�ĺ�����ʹ�����м����Ĥ��ص�֬Ƥ��1��2Ũ�����ߡ�֬Ƥ�ؾ��н�ǿ�Ŀ�����������ЧӦ��������PAF�������ȵĺϳɣ���ģ��GC���ڶ����á������ʪ�Խ����ߣ�֬Ƥ��1��������Ĵ���GC�ķ�Ӧ��ȱ���йء�
(8)GC��ճ�����ӱ���ĵ������ã�GC����һ���ߵ��ػ����漰ELAM��ICAM��GC������Ѫ����Ƥϸ������ELAM-1��ICAM-1mRNA�͵����ӣ������ÿɱ�GR����RU486����ϡ�GC�ɵ��ڰ�ϸ����ѭ�������������߷�Ӧ���Ӷ�������ǿ��Ŀ�����������ЧӦ��
(9)GC��ijЩø�ϳɵĵ������ã�GC����ǿ�����Ƴ���άϸ���ϳɽ�ԭø�����ƶ�����֬ø�ĺϳɣ�Ӱ��ijЩ��������ø�ı���ٽ�Ѫ�ܽ�����ת��ø(angiotensin-converting enzyme,ACE)��������������ø(neutralendopeptidase,NEP)�����ɣ�ACE��NEP�ɷֱ���ļ��ټ��ĵ���֢���ʡ����������᳦��֢�Լ���ʱ���ֲ���֯���ټ�������NK1�������ߣ����ɽ鵼SP�����ߵ��ڼ��������á�������ı�������GC�����ơ�
(10)GC��MO�����ĵ������ã�NO��Ϊϸ���ڼ�ϸ������Ϣ����Ҳ�������߷�Ӧ����ٽ���֢��֯��Ѫ����������NO�ϳɵ�øΪNO�ϳ�ø(NOS)���������ϸ�����ӵĵ��ء�����֤��GC������NOS�Ļ���ת¼���Ӷ�����NO�ĺϳɺ��ͷš�
Ŀǰ���֣� GC�����߹��ܵĵ��ز������������Եģ���ijЩʵ�������£������ϼ�����GC����ǿ�ܰ�ϸ������ֳ��Ӧ��
����������GC�����û��������˷���ˮƽ����ʶ��GC���������GR��ʹGSP90��immunophilin�ȵ�����GR���룬�л��Ե�GR����ʶ����Ƥ�ʼ��ط�ӦԪ��(GRE)���У�����GREλ��GC�л�������������ʿ��յ������ְл���ı��GRE����Ŀ�������ת¼��ʼ�ϵ��λ�ÿ��ܾ���GCӰ��ת¼�ij̶ȣ�GC����ijЩ�л���ת¼�Ļ��ƿ��ܺ�������GRE�йأ�GR�����GRE��Ϻ��Կռ�λ�跽ʽ��������ת¼���ӵĴ�ת¼���á��½�������GR����ֱ��������ijЩת¼���ӣ��Ӷ����ӱ���ЧӦ����ת����AP-1(�����-1)����Fos����Jun������ɵĶ����塣AP-1�ٽ���ԭø����Ļ���л��Ե�GR�ɽ����AP-1�Ӷ����ƽ�ԭø�ĺϳɡ�AP-1������Tϸ���Ļ���ٽ�IL-2��IL-2R����ı���ȣ�GC��ͨ��GR�Ļ��AP-1�Ļ����ԶԿ�ϸ�����ӵ����á�����ת¼������NF-��B��NF-AT��Ҳ��GC�ĸ��Ե��ء���������о����Ժ�����ҩ����Ƽ���ص��ٴ�ʵ������ָ�����塣
2.������ һ����ԣ�غͪ���ۼ����������߹��������������á�غͪ�ɼ�����������IgA�IJ�������һ����Ϊ�ۼ��������С���غͪ�������£����ٵ���������������١�С��ٷ���Ƥ��������Ӧ���������������غͪ�����ƣ�ȥ�ƿ���ת��Щ�仯��غͪ���ɽ���ʵ�鶯�������ϸ����֧ԭ�塢�����漰������Ⱦ�ĵֿ���������Ϊ��Ⱦ��������ʼ������������������ߣ�ȥ��Ϊ������ת��Щ�ı䡣
����ͪ��E2�෴��غͪ����������Ƥϸ������Ia���ӡ����⣬IM-9ϵ�ܰ�ϸ��Ĥ��SP����(NK1����)��غͪ��������Kdֵ������ʾغͪ��SP��һ��Դ����֢���ʵ�������һ��Ӱ�졣
3.�Ƽ��� �Ƽ��ؿ������Һ������������ϸ�������ܡ�
(1)����Һ���߹��ܵ�Ӱ�죺E2�ٽ��ӹ�����IgG��Ҳʹ�ӹ���Ĥ��Ƥϸ����IgA�������ӡ�C57BLС����ܴƼ��غ����SRBC����Ѫ�Կ���ζ���������C3HС���˷�Ӧ����ʾ�����ô���Ʒϵ���졣
(2)��ϸ�����߹��ܵ�Ӱ�죺�Ƽ����Ƽ��ܽ������������������������ܰ�ϸ����Ŀ����������Ƣ���������Ƣϸ���������������£�����С���Ƣϸ����ĿҲ������Ϊ�ࡣ��ȥ�ѳ��Ĵ��Դ���E2����������ή����E2����ǿ�ҵ�����PHA��ConA�ȸ߲�Ĵ�������ϸ����ֳ��Ӧ��E2����С���˵�����ѪT�ܰ�ϸ��DNA�ĺϳɣ�E2������NKϸ���Ļ��ԣ�����poly I-C����С��״�ž���NKϸ���Ĵ̼����ã�ͬʱ����С��ʵ���Ի��Է��������ķ����ʼ�ת���ʣ��ӳ�ͬ������Ƥ����ֲ����С��Ĵ��ʱ�䡣ȥ����������PRL�ɻָ����߹��ܣ���ͬʱ�г��ѳ���PRL��������ȫ�ָ����߹��ܣ��������E2������Ч��FE������߷ʴ�ϸ������Ŀ���̼��ʴ�ϸ���Ĺ��ܡ�
4.ȩ��ͪ ȩ�̱����Ĥ���������Ӱ���˵����˰�ϸ����Na+�������Ӷ��ı�ϸ���ڵĸ��ӳ��Լ�ϸ�����������ƿ�����IP3�������йء�
��״�ټ��ض���Һ���ߺ�ϸ�����߾��дٽ����á��������������ȥ��״�ٺ���������Ѫ�ܰ�ϸ����Ŀ���ͣ���SRBC�Ŀ��巴Ӧ�½���Ƣϸ����PHA�̼�����ֳ��Ӧ��������ЩЧӦ��һ����ʱ�������ԣ�����������ȥ��״�ٺ�������仯�����ڶ��������������辭40-60�������Ƹı䡣�ɼ�����״�ٶ������������Ե������á��Ŵ�������ȱ���Snell-BaggС�������������(GH)��T4����ؽ������߹��ܡ�T3������������С��������Ƥϸ����Ŀ��������������������ڸ���T4���������Ѫ�ܰ�ϸ���������ر���Tϸ����Ŀ����С���ˣ���״�ټ��ؾ��ٽ��ܰ�ϸ����˿��ԭ����ֳ��Ӧ���������Եļ���������ϵ��
1.GH �˵�GH��191��������л���ɣ���ͬ������GH�ṹ�ͻ����нϴ�IJ��졣GH�������弤���м���Ҫ�����ߵ������ӡ�GH�����Ǻ�ϸ��������(EPO)���峬�����һԱ��GH�ȿɽ�������ֱ��Ӱ������ϸ���Ĺ��ܣ�Ҳ�����ȵ�������������(insulin-like growth factor,IGF)�Ľ鵼���������������ϸ������GH�յ�IGF�����ɣ���IGF-I����ֲ�����������Ѫ������ϸ��Ĥ�ϡ�GH��Ӱ������ϵͳ�ĸ������ڣ���������Ϊ����Ҫ�İ����١�
(1)С��ȥ���������������������٣��ܰ���֯ή����DNA�ϳɼ��٣��ɿ�ԭ�շ��Ŀ��巴Ӧ������Ƣ����NKϸ�������½�������ȥ���������Ϊ���ٺ��ܰͽ����ֳ��Ӧ������Ƣ����DNA��л��������Ƥ����ֲ���ųⷴӦ�����ƣ������շ������Թؽ��ף��ҿ���ϳ����������GH�����תȥ�����������仯��
(2)���������幦�ܵ�������٪��֢������ƶѪ��EPO���͡���GH������ٽ����������ӹ������ܰ�ϸ������Ŀ������Ѫ������EPO����Ҳ���ߡ�
(3)�����Ŵ��Դ�����٪��С��(Snell-BaggС��)�������ܽϲͻ������Ϊ�����˻��磬�����ܰ���֯��������ϸ��ϡ�٣����߷�Ӧ�Խ��͵ȡ���GH��T4���ϴ�������С�ɷ�ֹ����ή��������ܰ�ϸ����Ŀ���ָ������ܡ�GH����һ���ÿ����������ٽ鵼�ģ���ȥ���ٺ�GH�ٽ����߹��ܵ�������ʧ�����ѷ���GH�ܴٽ����ټ��ص��ͷš�
(4)˥��ʱ��GH���ڼ��٣����߹��ܽ��͡�����GH�Y�лָ����߹��ܣ��ٽ����ټ��صķ��ڣ�����Tϸ��ǰ��������٣�������˥��С�������������GH��GH3������ϸ����ֲҲ�������������������ϸ����Ŀ��
(5)GH�ٽ�������Tϸ��������γɣ����̼��ܰ�ϸ������ֳ��GH�ɼ�ǿPMA�Ե�����ϸ���ͷ�H2O2�Ĵ̼�ЧӦ��GH���ɼ�ǿ������ϸ�����������ԣ������ÿɱ�SS������M����GH�̼��¶Ե��ܶ�֬����(LDL)����ȡ�ͽ�����١��½�����֤��������GH��ֱ�Ӵ̼�Bϸ����ֳ������Ig��
����������ʵ�����������Ӧ��GH���ƹ��蹦��˥�ߡ����߹��ܵ����Լ���˥�ϡ�
2 .PRL PRL���������м��������ڡ��ڵ͵ȱ��¶��PRL������Ҫ�������ѹ�������������ʹ�л�ȵĵ��ڡ���һ�ز��鶯�P���࣬PRL����ֳ����������أ���̼����鶯�����ٵķ����ͷ�����֭�����Ը������ٵ������ͷ��ڣ����ٴ���Ļ������ɼ��ܽ�ȡ��ڷ��ӽṹ�ϣ�PRL��GH��̥�̴����ؾ��������ƣ���PRL��GH�ĸ�Ѫ����������¼���ں�ϸ�����������峬���塣ͬGH���ƣ�PRLҲ�����߹����������������á�
PRL�����ٽ����ٷ��������飬������������з���IgA��ϸ����Ŀ���ܰ�ϸ�����߽������١�ĸ����PRLŨ����Ӥ��Ѫ��IgG��T�ܰ�ϸ����Ŀ������أ�˵��ĸ��ι������Ҫ�����ߴ̼����á�ȥ���嶯������������ֲͬ�����崹��ɻָ��������߹��ܣ��翹SRBC�Ŀ����γɣ���DNCB�ĽӴ���ÿ�������Թؽ��ķ�������֪��ֲ������Ҫ���ڴ�����PRL����PRL��������ֲ����ָ�����������Ҫ���أ���Ϊ��������PRL�Y�лָ�ȥ���嶯��������ܡ���֮����ȥ���嶯��ע�俹PRL��Ѫ�壬��֧���ƶѪ��������������PRL���ڵ�ҩ��Ҳͬ�����ƶ����DNCB�ĽӴ��Գ�����Ӧ��
PRL�����ߵ��������漰���·��棺�ٴٽ�����ϳɣ��ڴ̼�������ϸ���ķ�����ֳ���۴ٽ�������֯���������յ���̥������ϸ������Thy-1��ԭ��Tϸ����ԭ������IL-2Эͬ�̼�Tϸ���Ŀ�¡��ֳ�����յ������IL-2R���Կ�¡�� Tϸ��L2��ϵ��PRLҲ��IL-2Эͬ�յ����������ص�������(IRF-1)��C-myc��ODC(��������ø)���鵰��cyclin B�ȷ��ӣ��ٽ�Tϸ������ֳ���ݴ̼�����Nb2�ܰ�������ֳ��������Dex�����DNA���ѣ���ֹ���Կ�Nb2ϸ����Dex���õĵ��㣻����M�գ�����NKϸ��������IL-2Эͬ�յ�LAK���ԡ�
3.ACTH ACTHΪ39�ģ���Ӱ���������ϸ����������ˮƽ��ACTH��ЧӦ���پ��ɶ���;������һ�Ǵ̼�GC�ķ��ڶ���������������ƣ�����ǽ�����������ϸ��Ĥ�ϵ����������ֱ��Ӱ�����߹��ܡ�ACTH���Ƽ���������ή����Ƣή���������ܰ�ϸ����Ŀ���٣���ȥ�����ٺ��ЧӦ��Ȼ���ڡ�
(1)ACTH��Bϸ�����ܵĵ��ڣ�ACTH�����������Tϸ�������Կ�ԭ(��SRBC)�����ź������Կ�ԭ(��DNP-Ficoll)�Ŀ��巴Ӧ������PFC��Ŀ�������þ������Ե�ACTH���ӽṹ�����ԣ���ACTH(1-39)Ϊ�����Եģ���ACTH(1-24)��Ч��ACTH������IL-2��DCGFЭͬ�̼�������Bϸ�������ͷֻ�����BCL1ϸ��ϵ���ԣ�ACTH�������Է��ڷ�ʽ�ٽ�����ֳ��
(2)ACTH��Tϸ�����ܵĵ��ڣ�ACTH����Tϸ������IFN-�ã�������IL-2�����ɡ��½����֣�Jurkatϸ��Ĥ��CD3���ӵĦ������ữ����Ҳ��ACTH�ĸ��Ե��ء�ACTH����ǿ����ܰ�ϸ����Ӧ(MLR)�е�ϸ�������á����⣬ACTH����ȫ��ֹIFN�յ�M�յ�ɱ���������ԣ������Ƹ�ǻM�յ�MHC������ӱ��
4.��-END ��-END��ACTH��Դ�ڹ�ͬ��ǰ��POMC�����ڦ�-END�����������ͷţ�Ҳ��������ϸ���кϳɣ���������㲼�ڶ�������ϸ�����ʦ�-END���й㷺�����ߵ������á�
(1)��-END��Tϸ����Ӱ�죺��-END�ɴٽ�Tϸ������ֳ��Ӧ����Ҳ���෴��������-END��������Tϸ������IL-2R������������ѪTϸ��(HPBTL)��õ�廨���γ��ʣ��ı�Tϸ��Ĥ��CD3ԭ�ı��Ӱ��PHA����CD3���������ữ�Լ�����CD3-TCR��������ڻ����̡�
(2)��-END����������ϸ����Ӱ�죺��-END�Լ���������ʽ�ٽ�NKϸ����ϸ�����������ɻM�մٽ������ɺ��������ԣ����ɵ���MHC ������ӵı��
����-END�⣬��-END����-ENDǿ����(dynorphin)��M-ENK��L-ENK�Ȱ�Ƭ������ɽ������ְ�Ƭ�������������ϸ���Ĺ��ܡ�
5.CRH ��Ϊ41�ĵ������Լ��أ�CRH���پ��ɶ���;��Ӱ�������ܣ�CRH�ɵ�������AVPЭͬ�̼�ACTH���ͷŶ�����GC�ķ��ڣ�����߾����й㷺����������ЧӦ��CRH��������ϸ��Ĥ�ϵ������ֱ��Ӱ������ϸ��������CRH�������ټ�Ƣ����������ٺϳɣ���CRH���ܾ�����Ҫ�����������ߵ������á�CRH����������Ѫ������ϸ������IL-1�¼�IL-6��CRH������������IL-1�µ����ɣ�����IL-6�̷��Է��ڼ��١�����ʱCRH��GC��Ѫ����Ũ�Ⱦ����ߣ��ʿ�������ĸ���̥�ҵ����߷�Ӧ��
Ҳ�����ױ�����CRH��������ǿ���ã�CRH�ɴ̼�������Ѫ������ϸ������IL-6������IFN-�õķ��ڣ�����Ӱ�����ϸ������ֳ��IL-1�µķ��ڡ��б���nMˮƽ��CRH�ܴ̼�������Ѫ������ϸ������IL-2��IL-1����ǿLPS��PHA�ֱ��IL-1��IL-2�Ĵٷ��ڷ�Ӧ���̼���-END�ķ��ڡ�CRH���ܴٽ��ܰ�ϸ������ֳ��IL-2R(Tac)�ı���ˮƽ�����ɵ���NKϸ���鵼��ɱ��ϸ�����á�
6.LHRH ������Ƣ�������CRH����mRNA��Ҳ��CRH���������Ҵ�������ֱ�ӷֲ��ڴ���С����ܰ�ϸ���ϡ�Ŀǰ��Ϊ�����ٿ�����˥�ϵ�ʱ�ˡ�����������������ʽ����Լ��ᣬͬʱ����CRH������Ŀ�ļ��٣�������CRHǿЧ������(LHRHA)����ת��Щ�ı䣬���ڵ��û����ConAʱ�ٽ�����ϸ������ֳ��Ӧ���̼�IL-2R�ı��LHRH-A��ҪӰ��CD4+��CD8+Tϸ����Ⱥ������ȥ��Ҳ���������Ʊ仯�����ٸ���LHRH-A���Эͬ���á�LHRH�����ڻ���������дٽ����߹��ܵ����á�
7.SP �ڴӶ������У�SP�����ߵ���ЧӦ�о���Ϊ�㷺�����롣SP��Ӱ�����е�����ϸ����
(1)SP��������Ѫ�ܰ�ϸ�������ã�SP�м��������ԵĴٽ��ܰ�ϸ����ֳЧӦ������ǿConA��PHA�Ĵ̼���Ӧ��SP�������������ƴ˷�Ӧ�����SP������Tϸ����˿��ԭ����˿��ԭ��SP���ɴ̼�С��Ƣ�ܰ�ϸ��������Peyer�Ͻἰ��ϵĤ�ܰͽ��ܰ�ϸ������ֳ��SP����N��Ƭ��SP1-4�ɷ�ֹ����Ӧ�����µ�����ή�����ڶ��������ĸ��壬SP��ǿ�������Ĵ�T�ܰ�ϸ����ֳЧӦ�����з��֣�SP����������鵼����������Tϸ����Ca2+Ũ�ȡ�
(2)SP��Bϸ�������ã�SP�ٽ�Bϸ������Ig�ı�����Ϊһ�¡�SP�̼���������С��Peyer�Ͻᡢ��ϵĤ�ܰͽἰƢ����Դ��Bϸ��IgA�ĺϳɷֱ�����Լ300%��50%��70%�����ڸ���SP1-7�պ�����������֯���ܰ�ϸ��������ʱIgA��IgM�ĺϳ��������ӣ�������Ϊ�����Եģ�������SP����������������Χ�ڡ�SP��IgA�ϳɵ�Ӱ�������ԣ���ʾSP������ڑu�����ߡ�
(3)SP�Ե���-����ϸ�������ã�SP���ܴٽ������ɺ����������ԣ��ֿɴ̼���İ���������Ӧ(oxidative burst)���ٽ����ֽ��ʵ��ͷţ�������DP���Խϵ͡���С��ǻM�ռ������ܵ���ϸ����SP��SP1-4���������ɹ�����ǿ��SP�̼��ูǻM�յ�������������ǿ������ϩ��Ĵ�л�����ɺͷų�O2-��H202��TXB2��PGE��6ͪPGF1����LTC4����ø��øADGase(��-D-2-������-2-��������������ø)�ȣ�SP�ٽ�������Ѫ����ϸ���ͷ�IL-1��IL-6��TNF-�����ٽ�IFN-�õĺϳ�����ڣ��յ�С��M��ϸ��ϵP388D1ϸ���ͷ�IL-1��SP�ɻ���M-CSFЭͬ�̼�С����赥��-����ĸϸ��ϵ����ֳ��Ӧ��ͨ����������SP��ӵص���M����Tϸ�����ʶ�𡢿�ԭ�ӹ�����ʵȹ��̡�
(4)SP��������ϸ������������ϸ�����ã�SP������Ũ�ȼ��ɴ̼��˶��κ˰�ϸ���������˶���������ǿC5a ���µ�������ϸ�������������˶�������ɱ�����ԡ�SP���ܴٽ�������ϸ��ճ����֧������Ƥϸ�����ʿ��ܲ���������IJ������̡����⣬SP��ͨ���ʴ�ϸ�����ٽ���ϸ���Ľ���SP����������ϸ����Ӱ�첻�����ˣ���SPͬ���ɷʴ�ϸ���ѿ�����������������ϸ�������������ߣ�ȱ���ʴ�ϸ����WBB6F2-W/W�ü�WCB6F2-SL/SL2ϵС���˷�Ӧ����������С���j����ֲͬ�ֹ���ϸ����ֲ�ע��С��ʴ�ϸ����Ƥ�¸���SP��������ò�λ����������ϸ����������Ӧ��
(5)SP�Էʴ�ϸ�������ã����˼�����ʵ�鶯���֤����SP�̼��ʴ�ϸ���ͷ���֯����ЧӦ��SP���ٽ�5-HT�Դ������ϸ���ķ��ڡ�SP�շ��鰷��5-HT�ͷŷ�Ӧʮ��Ѹ�٣�30�����ͷ����ɴ����ֵ��90%�����������ǽͽ���������ữ���̡�SP����һ��������G���鵼�ģ�����Ҫ����Ca2+�Ĵ��ڣ�Ҳ������IP3��DAG�ĺ����仯����IgE�շ����鰷�ͷ���SP��ЧӦ�������������ʴ�ϸ���ܱ��P��������ϸ�����ӣ���TNF-����IL-1��IL-3��IL-4��IL-6��GM-CSF�ȣ�SP��ѡ���Եشٽ��ʴ�ϸ��ϵCFTL12����TNF-��mRNA���ͷ�TNT-�����������ñ���SP�Էʴ�ϸ����Ӱ������������ķ����ͷ�չ�����е���ϵ��
(6)SP������ϸ�������ã�SP�ٽ�����άϸ������Ĥϸ���Ƚ����֯ϸ������������ЭͬIL-1���³���άϸ����ֳ���ԣ����ɴ̼����ͷŽ�ԭø��PGE2����Щ��SP�շ��ʴ�ϸ�������鰷���Ӷ�����Ѫ�����š�Ѫ����ӿ������ϸ�������¾ֲ���Ѫˮ�ס��������Խ᳦�ס�ʮ��ָ���������ʪ�Թؽ������������ʵ���Թؽ��Ȳ��䲿λ��SP�����������ߣ���Ĥǻ��������Һ�м������ڶ����ݿ�ʱѪ��SPŨ��Ҳ��������Щ��ʵ����ʾSP��Ϊһ�����Խ��ʲ�����Ҫ�����߲������̡�
8.Ѫ�ܽ����آ�(A��) ��A������ߵ���ЧӦ�˽ⲻ�ࡣ��֤��A��̼�������Ѫ����ϸ������������Ӧ����ǿ�����������������Ӱ���仯ѧ���ԣ���Ũ��A��(10-9M)�������ɶ��տȶ���(PTX)���е�G���鵼�����������Ca2+Ũ������������Ũ�ȵ�A��(Ca-6M)����Ca2+�������٣�PLA2������ǿ���̼�������ϩ��Ĵ�л���ٽ�PKC����λ��
9.SS SS��ʹ�����������M�յİ������ǿ�������¶ȼ�Ca2+������ʽ�̼�����ǻ�ʴ�ϸ���ͷ��鰷��SS���ܶԿ�VIP����Ĵ����ܰ�ϸ���������ỷ��ø�������ӡ�
10.FSH(���ݴ̼���follicle-stimulatinghormone)��LH(���������أ�luteinizing hormone)������Ƥ�ʲ���LH��������ϸ���ķֲ����������ڲ�Ƥ�ʺ�������FSH��������ϸ���ķֲ�����ʾLH��/��FSH���ܲ�������ϸ���ķ������ܡ�
11.�ȵ��� ��С���ȵ��ؿɸı�ConA������ܰ�ϸ����ֳ��Ӧ����LPS����Ӧ�ÿ����ӿ����γ�ϸ����Ŀ���ȵ����ڲ��ı�Ѫ��Ũ�ȼ���ʱ�ܼ�ǿС��������ݿ˵ķ����������ƽDz�˽����µ��������͡������壬�ȵ��ؿɴٽ�MLR�е�DNA�ϳɣ�����ADCC���ܡ��������Ե��ȵ��ؼ��ɼ�ǿ����ϸ���������ԣ��ٽ���Ծ�ֹ���������ɣ�����߶��κ˰�ϸ�����������ԡ�
1.���豾�ְ� ��֧���ܰ����ٵ���ĩ���ͷŵ�ȥ����������(NE)�������������ͷŵ���������(Adr)��NE�����ɦ���������Ӱ���������ϸ�������߹��ܡ�����Ӱ������ø��Ӷ���������������ͬ������������ؿ�����С����̥����ϸ������Thy-1��ԭ��Adr�ɽ�����T�ܰ�ϸ����˿��ԭ�̼�����ֳ��Ӧ��������Һ����Ӧ�𣬵��¿���ϳɼ��ټ�I�ͳ�����Ӧ�����ơ�������ϸ��Ӱ����о������һ�£���Adr��NE������Ũ��ʱ����M�շ���IL-1�������б����Ʀ�2�����˷ܴٽ�M���ͷ�TNF��Adr��NE��������ϸ���������������ɻ��ԡ�����Ӱ���������ֲ�ųⷴӦ������GVHR���½����֣����豾�ְ�������������Ѫ����Ƥϸ�����ٽ�MHc I���������ӱ����NE���ڵ������ν���ϸ����MHC������ӵı��
2.ACh ������ҩ���簱��������(carbachol)������������ѪTϸ����E�����γ��ʣ��˷�Ӧ�ɱ�����Ʒ����ϣ�˵����M����IJ��롣Ach������M������ߴ���T�ܰ�ϸ����ϸ�������ã��ٽ�PHA���µ��ܰ�ϸ��ת���͵����ʺϳɡ��������˷���֯��ACh�̼��鰷���ͷţ���ЧӦ���ܰ���Ʒ�������ͼ�����AChֱ�Ӵ̼��ʴ�ϸ���ͷ��鰷��ACh���볦����Ӧ�ķ�������ţ���к���-���ף��������ţ��Ĺ�����Ӧ����-�������շ�����ĩ���ͷ�ACh����Ϊ������Դ�����߽��ʡ�
3.5-HT 5-HT������Ϊ���ʣ������ѪС�弰�ʴ�ϸ���ͷš�5-HT�ܽ��Tϸ����ֳ���������أ�Ӱ��NKϸ�����ԣ�����M�ձ���Ia���ӡ�
4.�ʺ��� �ɹ��������߹����������е���ϵ����ҩ������ɹ���Ĺ��ܡ�ժ���ɹ�������������յȴ�ʩ�����������ٳ��ο���ϳɷ�Ӧ�����ƻ���ܰ�ϸ����Ӧ���������ʺ��ؿ���ת��Щ�仯���ʺ��ػ��ɴٽ�С��Ƣϸ������IFN-�á�
����Ӧ��������ϵͳ��Ӱ��
(һ)�����Ӧ������
�����˺��Դ̼������һϵ�з�����Ķ��ͷ�Ӧ���ɳ�ΪӦ��������Ϊ������-����-������Ƥ����ϵ(HPa axis)��������-����������ϵͳ���˷ܣ��������ڶ���֯�����ٵĹ��ܱ仯��Ӧ������ȡ������绷�����صı䶯�����ʸ����������ص�Ӱ�졣
����Ӧ���Դ̼�����������أ��ɷ�Ϊ������Ϊ���ġ���������Ϊ���ļ����������(��10-3)��
��10-3 Ӧ��ʱѪ���м���ˮƽ�ı仯����
���ࣺ����Ӱ���NE��Adr,dopamine
CRH-ACTH-GC
��-END��HG��PRL��suppressin
ADH,RAA(renin-angiotensin-aldosterone)
NPY
PGSs,TXs,kinins
IL-1,IL-6
����insulin,LH,testosterone,TSH(����ʱ����)
ע�������������ʱ�(1991)
(��)Ӧ��������ϵͳ��Ӱ��
ʮ��������Ӧ��������ϵͳ��Ӱ�챶�����ӡ�����ʵ�鼰����۲�֤ʵ��Ӧ�������ߵ�Ӱ����Ҫ�������ԡ�
(1)�Ե����Ϊʹ��Ӧ���̼����ɽ��ʹ����ܰ�ϸ����PHA����ֳ��Ӧ����һ���������Ե���֯���죬��Ƣ���ܰ�ϸ�����Ե���̼�������Ӧ��������ɼ���Ƣ����NKϸ�����ԣ�����M������H2O2����Щ�ı���Ҫ���ɰ�Ƭ������鵼�ġ�
(2)��2-�Ѱ�������(2DG)����ϸ���������ǵ�������������Ϊ��л�Լ�ģ�ͣ�Ҳ������ƽ������Ӧ����Ӧ�ɼ���ʵ�������������Լ����ķ����ʼ������س̶ȡ�
(3)ʵ�鶯��������������ر���ÿ����������ֻ��������������Ӱ����������������ʹTϸ�������Կ������ɼ��٣�Tϸ��ת���ʽ��͡�����Ӱ��������������Ա���죬��������Ӱ��Ϊһ���ԣ�����ҪӰ��Thϸ����IL-2�����ɣ��Ҳ������������ٵĵĴ�����������
(4)Ӧ������С���HSV��Ⱦ��������������Ƣ����ϸ�������ܰ�ϸ����Ŀ������С��ʵ���������ĵ�ת���ʡ�
(5)�������������Ϊһ�ֵ��͵�Ӧ���̼����ɵ���Ѫ���м��غ�ϸ������Ũ�ȵı仯����GH��T4��IL-2�Ƚ��ͣ���GC��IL-6���ߣ���Щ�仯�ɱ�PG�ϳ���ϼ�����ʹ����ϣ��ܰ�ϸ����PHA�ķ�Ӧ���͡���ͯ�����˺�Ѫ���ܰ�ϸ�����٣���Th��Ts�ļ���Ϊ�����ԣ�ͬʱ��ϸ�������������½���IgM���Ͷ�IgA��������������ȱ����������������Ѫ�ܰ�ϸ�����࣬CD16+ϸ�����ӣ�NKϸ�����Լ��ܴ̼���Ļ��Ծ�������������ϸ����Ŀ�ļ��١�
(6)�������ض������߹��ܵ�Ӱ���Ϊ��������ۿ����������Ӱ���ɱ���Ӧ�����ɽ����ܰ�ϸ����ConA�ķ�Ӧ������Bϸ����PWM�̼�����ֳ��Ӧ�����Խ��н���������Ϊ����Ӧ��ʱ�������ֳ��ܰ�ϸ����ConA���µ���ֳ��Ӧ�½����¶���ǿ�����ǵȸ�����������������Ӱ�����ԭ���ܰ�ϸ���Ĵ���ֳ��Ӧ��ͬʱIL-2R����ˮƽ�½�������ѹ�����������͵������Ӧ���̼�������Ѫ�п�HSV����EBV��ϸ�������Ŀ���ζ�������CD4+ϸ����NKϸ���İٷֱ��ʼ�����Ҳ��Ӧ���ͣ���ʾӦ�����ܽ�����������ʹ����DZ������������⣬�����������߹���ʧ�����ǹ��ϵġ�
(7)������Ӧ�����������༲���ķ����ͷ�չ��ϵ���С����ڻع��Ի�ǰհ���Ե����о��з��֣���ͯ������Crohn's�������ʪ�Թؽ��ס�������Ĥ�ס�Graves�����Ϻ�������Ⱦ�ȼ������������Ӧ���¼��в�ͬ�̶ȵĹ�������Щ����Ӧ���¼��У�ͻ����������ȥ��������ʧҵ�������ʡ�������Ӧ��Ҳ����������������ʺ�ת���ʡ�
��Ӧ���Ŀɿ��ƻ�Ԥ��������Ч�ظ��ƻ���Ӧ����Ӧ������Ӧ���̼������������ƣ����ϳ���Ӧ��������������ǿ����������ΪӦ������ʱ��С��NKϸ�����ԡ��ܰ�ϸ����PHA�ȵ���ֳ��Ӧ������ˡ��ºͼ��Ե�Ӧ��ѵ����������ܰ�ϸ��ת��ˮƽ��
(��)Ӧ��Ӱ�����߹��ܵĻ���
����Ӧ���������߹��ܸı�Ļ�����Ϊ���ӣ��������¡�
(1)CRG-ACTH-GC��ϵͳ�ļ��
(2)����-����������ϵͳ�ļ��
(3)��Ƭ����IJ����磬��-END��M-ENK��L-ENK��Dynorphin�ȡ�
(4)GH��PRL����-MSH�������ӣ�LH��FSH��TSH(��̼�����)�ȷ��ڼ��٣���-MSH�ɶԿ�ϸ�����Ӽ���HPA��ϵ�����á�
(5)IL-1��IL-6�ȵ��ͷš�
(6)����ǰҶ���ɵ�suppressin(����������)��Ϊ63kDa�ĵ����ʣ����н�ǿ����������ЧӦ����ACTH��GH��PRL�ȴ̼�ϸ�����ɣ��ʿ��Ʋ���ΪӦ���Լ��أ���������߹��ܵ������Ե��ء�
(7)Ӧ��ʱѪ���г��ֶ��������������ӡ������������ˡ�ʧѪ�Ⱦ��̼��������ɶ���Ѫ�������������ӣ��������ܰ�ϸ������IL-2��GC��PGE2���ء��Ǵ�����Ӧ��Ҳ������Ѫ���г���һ�������������ӣ�������Ϊ���ȵ�6kDa�Ķ��ģ�����������С���ܰͽ��ܰ�ϸ������ֳ�������ӵ�������������ϵͳ�IJ��룬�����漰�������ؼ���-END��
����Ӧ��ʱ�ڶ����ڷ��ڹ��ܱ仯�����Զ���;����ˮƽ�ı��������������ر��ǽ���Ѫ�������������Ӽ��ٴ����suppressin�ķ��֣��������ڲ���Ӧ��ʱ�������ڷ�������õı仯���ɼ������������塣��Ӧ��ʱ���������ƿɱ����������ܸ����ص����ˣ�����һ����ȴ���ͻ���Բ�ԭ��ĵֿ����������������������Ⱦ�������ķ�����
�ġ���ϵͳ��λ��ٶ����߹��ܵ�Ӱ��
(һ)�����ṹ��ٵ�ЧӦ
�����й�������ϵͳӰ�����ߵĹ�����Ҫ�����ں��ż���·��ټ�����������١������и��ѧ��ٵȷ�������1958�꼴�۲쵽�����Խ�ڲ����ƻ��ɸı���������߷�Ӧ�ķ����ͷ�չ���Ժ��о���һ�����֣�������ϵͳ�����ಿλ���˺�������������ܱ仯���������������������٣�����������ЧӦ����ʾ��Щ����Ϊ���߷�Ӧ��ǿ�������������ǰ����Ƣϸ��������ϸ������Ŀ���٣���ԭ��˿��ԭ�̼����µ��ܰ�ϸ����ֳ��Ӧ������������ʵ���Ա�̬��Ӧ�����ס�С�������Ը��ڲಿ�����ڲಿ����״�˵��ƻ�����NKϸ�����Խ��͡����ݶ���Ľ��©�������˿ɴٽ������������ڱ�Եϵͳ��������ٵĽ����Ϊ������ǿЧӦ������ͷ�塢�����м����ʺ˵��ƻ�������Ϊ����ܰ�ϸ������ֳˮƽ�������ƻ�����������ϸ����Ŀ���˴���Ƥ�������������ܱ仯����ijЩ�ص㣬�����Ƥ�����˰���Ƣϸ����Ŀ���ͣ��ܰ�ϸ����ֳ��Ӧ������NKϸ�������½������Ҳ�Ƥ���������������ı䣬��ʾ����Ƥ���������ܵ����ϵķֹ�������ٴ��۲�˵�����������������Լ���Ҳ֧������ʵ���������ڵ���Ƥ�����˺�����߱仯������3�����ˣ�����ʾ������ij�ִ����Ի��ƵĴ��ڡ������Բ����г�����ٿ�����������Ƥϸ���ļ��٣���ʾ�����v�����ڣ���ϵͳ������ϵͳ�伴�й��ܵ���ϵ��
���Ϲ���Ҳ����һЩ�ؽ����⡣��һ���ӷ���ѧ�Ƕȿ��ǣ�����ṹ������������߶Ⱦ���ȷ�����꼰����ṹ��ͨ·�������������״̬�ı������Ϊ�Dz�λ����������ġ��������ЩЧӦ�ķ���;�������ƾ�������в���������������ϱ��������Ե�ˮ�ܼ�ӱ����ٺ��������ǿЧӦϵ�ɸ����������ٵ�ֱ��֧�����鵼�ģ��������ЧӦ���漰GC���������弤���йء�
(��)��������ٵ�ЧӦ
Ƣ���г���С���SRBC�̼��Ŀ��巴Ӧ��ǿ�����������궯���������6-�Ƕ��(6-OH-OA)��ѡ�����ƻ����ܽ��������ʹTϸ���������Կ�ԭ�̼����������Ӧ����ǿ��
�г����߶��ﵥ��ĵڶ����������ܼ�ǿͬ���������ܰͽ��PFC��Ŀ���æ�-���ᰱ�����������������ʼ��صĺϳ���ɽ�һ����ǿ�˷�Ӧ��˵��������ȥ������������ϵͳ�����ߵĽ���������ЧӦ�����ܽ��������ߵ�������������������죬����С��Խ�����������������߱仯�ϴ�������ԡ�
�塢������������߹��ܵ�Ӱ��
(һ)����������������о��Ź�
����ѧ�ߵ������о������ѷ��ֿ��������Ӧ������ʽ�������䡣��70�������������������ҩ�ﻷ������Ϊ�������Դ̼�(unconditionedstimulus, US)���������Ǿ�ˮ��ζ���̼���Ϊ�����̼�(conditioned stimulus,CS)��ͬʱ�������������ִ̼�������һ��ʱ���(3��)���ٵ�������CS�����������Ե��������ƣ�����ΪTϸ�������Կ���(��SRBC)�ϳɼ��٣�����������˷�Ӧ��˵���ѽ������ܸı�����Ӧ�����Ϊʽ�������䡣�Ժ���о�������ͬ���ɽ������ϸ�����ߵ��������䣬���������ӳ������������Լ���(����Ǵ�)С������������⣬��������Ҳ�ɵ���������ǿЧӦ�������NKϸ�����Եȡ���10-4��������Ϊʽ�����������������Ӧ��ı䡣
��10-4 ��Ϊʽ�����������������ЧӦ�仯
�����̼�
��������
���߱仯
�
��������
���SRBC�Ŀ��巴Ӧ��
��TNP-LPS�����
�������˾������
GVH��
�ӳ������������Լ���С�������
�����Թؽ��ײ������
ƢNKϸ�����ԡ�
���ܰ�ϸ��Ѫ�塢��
����ܰ�ϸ����Ӧ����
������
Ѫ��CD8+��
�� ��
��ԭ(BSA+����)
��ԭ�������֯���ͷš�
polyI:C
NKϸ�����ԡ�
�������ֿ��ԡ�
���+�ƹ�(��ֲ����)
ͬ������Ƥ����ֲ��
ϸ������T�ܰ�ϸ��ǰ���
���ԣ�Grossmann,Hervberman��Livnat(1992)
(��)��������������ķ���������˵
�����̼��������߹��ܸı�Ļ����в������GCŨ��������������ȫ�����������䵼�µ���������ЧӦ����Ϊ��US�̼�������GC����ʱ�����ı�����Ӧ��Ŀǰ������ѧ˵����������������ߵ��ڻ�����
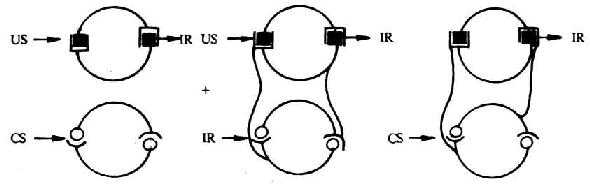
1.���ڹ�����ѧϰģʽ(associative brain learning paradigm) ��ѧ˵��ΪUS�����������Ϣ��CS��Ϣ��Ϊ��������źţ��ɼ���������ͨ·������������������·�佨����ǿ����ص���ϵ(�ṹ�Ĺ����Ե�)���ú�����CSʱ����������·��������ɴ������������ϵͳ��ѡ�����źţ���������Ӧ������������ã���ͼ10-3��ʾ���¡�
ͼ1-3 ���ڹ�����ѧϰ����ģʽ
����Grossmann,Heberman��Livnat��(1992)��
ע��US:�������̼� CS�������̼� IR������Ӧ��
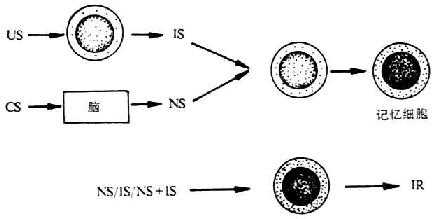
2.����ϵͳ�ڲ��Ĺ�����ѧϰģʽ (associative learning within the immunesystem) ����������ǽ�CS�Ĺ�����ѧϰ���̶�λ������ϵͳ�ڲ�������ԭ��ҩ��̼��ܰ�ϸ��ʱ��ijЩ�ܰ�ϸ�����ɴ����ɵĸ����źŵ���������ǿ��ͬʱ������ϵͳ�������̼�������Ӧ���������ܼ��ػ���ʵ��ͷ����ࡣ������������ܰ�ϸ����ͬʱ�������źŷ�����Ӧ�����ж��ͻ�����Ӧ��ļ��书�ܡ���������CS�̼���CS������Ӧ����Դ������źţ���������Щ����ʶ��ϸ�����ظ��̼���Ϣ�����´���ϸ������������ͻ�����Ӧ����ͬʱ����CS��US��������Ӧ��ϵ�������CSʱ��Ϊ���ԡ�������������߹��ܵ��ڵ�ʵ����ͬʱ������ܰ�ϸ������ϸ����ö�CS��US˫���źŷ���ʶ�ǺͶ��ͷ�Ӧ����������ͼ10-4��ʾ��
(��)�������������䷢��;��ʵ�����
Ŀǰ������֤��֧��ǰ����һ��˵������������ζΪCS����Poly I-CΪUS����С�������������䣬�������Ƣ����NKϸ���������ߡ��Ժ���CS��ɹ۲쵽NKϸ���Ļ�����������һ����������һ��������ķ������Ƽ�;����ȡ��������ʶ��
(1)����Poly I-C��ģ�ⲡ����Ⱦ�����Դٽ�IFN-�µķ��ڣ���˵���IFN-�¿�ȡ��Poly I-C��ΪUS��
ͼ10-4 ����ϵͳ�ڵĹ�����ѧϰģʽ
ע��CS�������̼� US���������̼� IS�������ź�
NS����ϵ�� IR���ⷴӦ��
(2)Ϊ̽��IFN-������β��뷴�佨���ģ�������׳�ע��100U��IFN-��Ҳ����Ƣ����NKϸ��������������ͬ������IFN-�¾�ע��Ч����IFN-�¿���ע������׳أ��������������Ľ���������Ӱ��US����ķ�Ӧ��
(3)US�Լ��������Է�ʽӰ�����߹��ܣ���PolyI-C 10��g/С��������Ƣ���NKϸ�����ԣ��������ɵ�IFN-���٣������Խ���������Ӧ��PolyI-c 20��g/С����������Ƣ����IFN-�¡�IFN-������mRNA����IFN-��ע��������ΪUSʱ����������10000U����Ч����������׳���ע��100U��IFN-�¼�����Ч�������PolyI-CΪUS���н��ź�ΪIFN-�£����źſ��ϴ������ڡ�
(4)CSΪ����̼���USΪ����������ǿ��OolyI-C���˶����ź����ϵIJ�λ�����������ԡ��ѷ�����������Ľ����ܶ������ص�Ӱ�죺Ԥ�ȸ�������ͪ��ϰ�Ƭ�ĵ�ЧӦҲ���������Ӧ�ij��֣����ò���Ѫ�����ϵİ�Ƭ������ϼ�QNTX����Ч����ʾCS����NKϸ������������������������谢Ƭ�ĵIJ��룬�ҿ���������������λ��
(5)��ҩ��Ľ��������Ӱ����ɷ�ֹ������Ӧ�ķ�������ʾ�����ڶ���Ӱ�������������Ľ��������֡�
�������˵������������Ľ����漰���ּ��䣬��һΪ���������ϵͳ�е�CS���䣬���ΪCS��US�������䣬�����ڸо�ϵͳ��������ಿλ���п����������ԡ������һ��ѧ˵���Ǻϡ�