第二节 免疫网络学说
免疫网络学说最早由Jene提出,在免疫网络学说的发展过程中不断得到完善和发展。
一、Jerne的免疫网络学说
Jerne强调免疫系统中各个细胞克隆不是处于一种独立状态,而是通过自我识别、相互刺激和相互特约构成一个动态平衡的网络结构。构成相互刺激和相互特约的物质基础是独特型和抗独特型。
1.Jerne的免疫网络结构 见9-5示意图。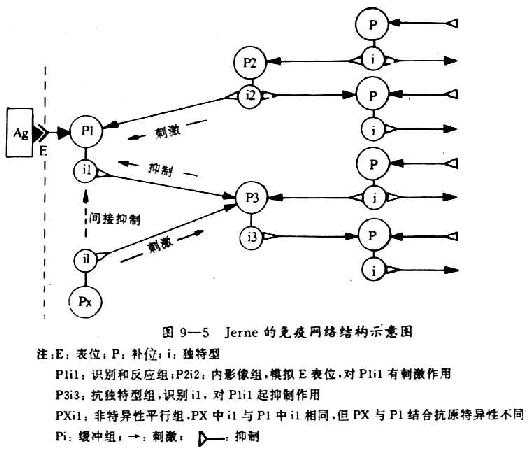
2.Jerne免疫网络解释免疫应答
(1)启动免疫应答:正常生理情况下,体内i2对P1的正调节和P3对il的负调节,使Plil处于一个动态抑制性稳定状态。外来抗原(E)进入机体后打破了识别和反应组、内影像组、抗独特型组以及非特异平行组之间保持的抑制性平衡状态。进入机体的抗原,一部分刺激识别组细胞,一部分立即与识别组预先产生的自然抗体相结合并消除这部分抗体,这就暂时减弱了识别组对内影像组的抑制效应和对抗独特型组的刺激效应。由于内影像组抑制效应的减弱,使该组细胞发生增殖并产生抗体,增强了内影像组对识别组的刺激效应。同时识别组对抗独特型组的刺激效应减弱,使独特型组处于更可抑制的状态,以致减弱了对识别组的抑制效应。因此,抗原的刺激、内影像组刺激效应的增强和抗独特型组抑制效应的增强等多种因素促进了识别组的增殖和抗体的分泌。
(2)免疫应答的自控:由于抗原启动免疫应答,识别组分泌抗体增加,恢复了对内影像组的抑制效应和对抗独特型组的刺激效应,使免疫应答水平得到控制,免疫网络的动态趋于恢复平衡。如刺激的抗原在体内持续存在,对识别组的刺激效应持续超过对该组的抑制效应,可使免疫应答持续发生。当抗原被清除后,对识别组的抑制效应超过对其的刺激效应,使免疫应答恢复到原先的平衡状态。
图9-6是简化的Jerne免疫网络结构示意图。机体的免疫网络同四组淋巴细胞构成。第一组为抗原反应细胞(ARC),可通过其抗原受体对外来抗原起反应,并以ARC为主体与另外三组淋巴细胞构成网络。第二组为独特型反应细胞(IRC)或独特型组(idiotype set)。能识别ARC上的独特型决定簇,从而抑制ARC对外来抗原起反应。第三组为内部影像组(internal image set)。其抗原受体上的独特型与外来抗原相同,故能被ARC识别,激发ARC增殖。第四组为非特异性平行组(nonspecific parallel set),它的抗原ARC不同,但其抗原受体上的独特型(Id)与ARC上的Id相同,故能被IRC识别,从而间接抑制ARC的增殖,加强了对网络的抑制作用。二、三、四组淋巴细胞又可各为主体,再分别与另外的三组淋巴细胞组成网络,如此下去,在体内形成网络系统。从总的效应来看,是一种均衡性抑制。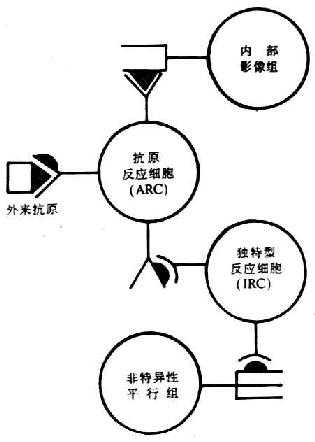
图9-6 免疫网络学说(Jerne)
免疫网络学说和内影像可解释医学上的一些现象,如记忆淋巴细胞的存在、Weigle现象以及非特异性的反应等。
(1)记忆淋巴细胞:Ab2β不仅可激发针对外来抗原或自身抗原Ab1产生前体细胞(precursors),使机体对入侵到体内的抗原迅速发生免疫应答,而且与记忆细胞的存在有关。当抗原被其诱发的免疫清除后,由于记忆细胞识别抗原受体具有更高的亲和力,因而Ab2β替代原始抗原可能足够刺激记忆细胞的增殖和存活。
(2)Weigle现象:现已发现,与自身成份有交叉反应的抗原能够使机体原有的免疫耐受丧失,这与某些自身免疫病的发生有关。例如,链球菌与心肌组织有共同抗原,正常情况下,机体对自身的心肌组织存在着免疫耐受。某些病人感染链球菌后,由于足够量的交叉反应性抗原刺激,增强了内影像组对ARC组的刺激效应,减弱了IRC组对ARC组的抑制效应,致使ARC组逃脱抑制,耐受性丧失,从而发生风湿性心肌炎。
(3)非特异性反应:某些抗原刺激机体后,可诱导出现非特异性抗体应答。Oudin发现,这一非特异性抗体与特异性抗体的独特型是相同的。依照网络学说,当抗原的刺激使ARC组增殖并分泌抗体时,加强了对IRC组的刺激,继而IRC组的增殖又加强了对非特异性平行组的刺激并使后者增殖和抗体分泌,这些抗全具有与ARC相同的Id,但结合抗原的特异性与ARC不同,即所谓非特异性抗体。
二、免疫网络的其它模型
Jerne的网络学说奠定的用整体的、联系的观点解释免疫调节和免疫现象的基本思想。以此免疫学说为基础,Richter、Hoffmann等又加以修改补充提出了新的网络模型。
Richter把各种不同的克隆称为功能单位,以Ab0、Ab1、Ab2、Ab3等表示,每一个克隆包括T细胞、B细胞、抗体分子及T细胞因子。Ab1识别外源性抗原决定基,Ab2识别Ab1的Id,Ab3识别Ab2的Id,以此类推。Ab1除能识别外源性抗原外,还能识别该抗原的网络中的内影像,即Ab0。在网络内部,Ab0刺激Ab1,Ab1刺激Ab2,Ab2刺激Ab3……;反之,Ab3抑制Ab2,Ab2抑制Ab1,Ab1抑制Ab0。其中,刺激效尖所需要的抗体分子浓度高于抑制效应所需之浓度。这种刺激与抑制的相互调节使免疫网络处于相对稳定的状态。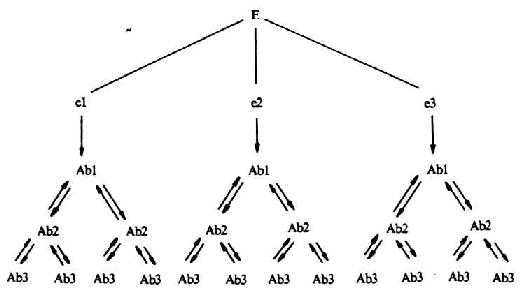
图9-7 Richter网络模型
按照Richter模型,可对免疫应答的抗原剂量信赖性反应进行解释。
1.正常免疫应答 适量的抗原刺激进入机体后,刺激相应的B细胞克隆产生Ab1,Ab1带有与产生Ab1B细胞表面BCR相同的独特型,当Ab1产生到一定水平时,可刺激体内识别Ab1上Id的另一组B细胞克隆产生Ab2;以类似的方式Ab2刺激机体产生Ab3。由于适量抗原刺激,反应只能使Ab3产生一定水平,此水平足以抑制Ab2,但尚不能刺激Ab4。这样,使Ab1克隆细胞得以活化增殖,产生较高水平的针对入侵抗原的抗体(Ab1)。
2.低剂量耐受性 低剂量抗原刺激机体产生较低水平的Ab1,只能微弱地刺激产生Ab2,其浓度恰好能够抑制Ab1但不足刺激Ab3,因此发生低剂量免疫耐受。
3.高剂量耐受性 高剂量抗原的刺激信号强,可产生较高水平的Ab3,进而刺激产生Ab4,Ab4对Ab3的反馈抑制,导致Ab2逃脱Ab3的抑制而增殖,从而抑制了Ab1克隆表现为高剂量耐受性。
Id除分布于Ig分子或BCR V区外,还存在于T细胞受体(TCR)α和β链,包括t h、Ts、TDTH和Tc等不同T细胞亚群。TCR上的Id可用抗IdAb或抗IdT细胞来鉴定。一般来说,TCR上的Id与Ig或BCR上的Id有所差别,这可能是TCRα、β链的V基因与Ig重链、轻链V基因同源性较低的原因。抗TCr Id也能模拟抗原而活化T细胞。最近研究表明,抗原特异性B细胞上Id不仅可被Ab2B细胞所识别,也可被独特型特异性Th、Ts或Tc所识别;同样抗原特异性T细胞上Id可被TCR特异性T细胞和Ab2 B细胞所识别,这可能与淋巴细胞V基因库中,TCRα、β链V基因库与BCR免疫球蛋白重量链和轻链V基因库有一定程度的同源性有关。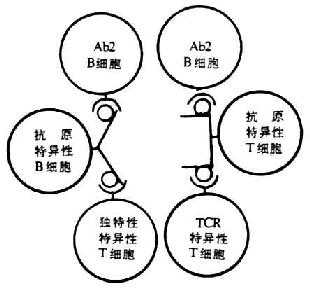
图9-8 TCR和BCR上Id相互被识别的示意图
一、Jerne的免疫网络学说
Jerne强调免疫系统中各个细胞克隆不是处于一种独立状态,而是通过自我识别、相互刺激和相互特约构成一个动态平衡的网络结构。构成相互刺激和相互特约的物质基础是独特型和抗独特型。
1.Jerne的免疫网络结构 见9-5示意图。
2.Jerne免疫网络解释免疫应答
(1)启动免疫应答:正常生理情况下,体内i2对P1的正调节和P3对il的负调节,使Plil处于一个动态抑制性稳定状态。外来抗原(E)进入机体后打破了识别和反应组、内影像组、抗独特型组以及非特异平行组之间保持的抑制性平衡状态。进入机体的抗原,一部分刺激识别组细胞,一部分立即与识别组预先产生的自然抗体相结合并消除这部分抗体,这就暂时减弱了识别组对内影像组的抑制效应和对抗独特型组的刺激效应。由于内影像组抑制效应的减弱,使该组细胞发生增殖并产生抗体,增强了内影像组对识别组的刺激效应。同时识别组对抗独特型组的刺激效应减弱,使独特型组处于更可抑制的状态,以致减弱了对识别组的抑制效应。因此,抗原的刺激、内影像组刺激效应的增强和抗独特型组抑制效应的增强等多种因素促进了识别组的增殖和抗体的分泌。
(2)免疫应答的自控:由于抗原启动免疫应答,识别组分泌抗体增加,恢复了对内影像组的抑制效应和对抗独特型组的刺激效应,使免疫应答水平得到控制,免疫网络的动态趋于恢复平衡。如刺激的抗原在体内持续存在,对识别组的刺激效应持续超过对该组的抑制效应,可使免疫应答持续发生。当抗原被清除后,对识别组的抑制效应超过对其的刺激效应,使免疫应答恢复到原先的平衡状态。
图9-6是简化的Jerne免疫网络结构示意图。机体的免疫网络同四组淋巴细胞构成。第一组为抗原反应细胞(ARC),可通过其抗原受体对外来抗原起反应,并以ARC为主体与另外三组淋巴细胞构成网络。第二组为独特型反应细胞(IRC)或独特型组(idiotype set)。能识别ARC上的独特型决定簇,从而抑制ARC对外来抗原起反应。第三组为内部影像组(internal image set)。其抗原受体上的独特型与外来抗原相同,故能被ARC识别,激发ARC增殖。第四组为非特异性平行组(nonspecific parallel set),它的抗原ARC不同,但其抗原受体上的独特型(Id)与ARC上的Id相同,故能被IRC识别,从而间接抑制ARC的增殖,加强了对网络的抑制作用。二、三、四组淋巴细胞又可各为主体,再分别与另外的三组淋巴细胞组成网络,如此下去,在体内形成网络系统。从总的效应来看,是一种均衡性抑制。
图9-6 免疫网络学说(Jerne)
免疫网络学说和内影像可解释医学上的一些现象,如记忆淋巴细胞的存在、Weigle现象以及非特异性的反应等。
(1)记忆淋巴细胞:Ab2β不仅可激发针对外来抗原或自身抗原Ab1产生前体细胞(precursors),使机体对入侵到体内的抗原迅速发生免疫应答,而且与记忆细胞的存在有关。当抗原被其诱发的免疫清除后,由于记忆细胞识别抗原受体具有更高的亲和力,因而Ab2β替代原始抗原可能足够刺激记忆细胞的增殖和存活。
(2)Weigle现象:现已发现,与自身成份有交叉反应的抗原能够使机体原有的免疫耐受丧失,这与某些自身免疫病的发生有关。例如,链球菌与心肌组织有共同抗原,正常情况下,机体对自身的心肌组织存在着免疫耐受。某些病人感染链球菌后,由于足够量的交叉反应性抗原刺激,增强了内影像组对ARC组的刺激效应,减弱了IRC组对ARC组的抑制效应,致使ARC组逃脱抑制,耐受性丧失,从而发生风湿性心肌炎。
(3)非特异性反应:某些抗原刺激机体后,可诱导出现非特异性抗体应答。Oudin发现,这一非特异性抗体与特异性抗体的独特型是相同的。依照网络学说,当抗原的刺激使ARC组增殖并分泌抗体时,加强了对IRC组的刺激,继而IRC组的增殖又加强了对非特异性平行组的刺激并使后者增殖和抗体分泌,这些抗全具有与ARC相同的Id,但结合抗原的特异性与ARC不同,即所谓非特异性抗体。
二、免疫网络的其它模型
Jerne的网络学说奠定的用整体的、联系的观点解释免疫调节和免疫现象的基本思想。以此免疫学说为基础,Richter、Hoffmann等又加以修改补充提出了新的网络模型。
Richter把各种不同的克隆称为功能单位,以Ab0、Ab1、Ab2、Ab3等表示,每一个克隆包括T细胞、B细胞、抗体分子及T细胞因子。Ab1识别外源性抗原决定基,Ab2识别Ab1的Id,Ab3识别Ab2的Id,以此类推。Ab1除能识别外源性抗原外,还能识别该抗原的网络中的内影像,即Ab0。在网络内部,Ab0刺激Ab1,Ab1刺激Ab2,Ab2刺激Ab3……;反之,Ab3抑制Ab2,Ab2抑制Ab1,Ab1抑制Ab0。其中,刺激效尖所需要的抗体分子浓度高于抑制效应所需之浓度。这种刺激与抑制的相互调节使免疫网络处于相对稳定的状态。
图9-7 Richter网络模型
按照Richter模型,可对免疫应答的抗原剂量信赖性反应进行解释。
1.正常免疫应答 适量的抗原刺激进入机体后,刺激相应的B细胞克隆产生Ab1,Ab1带有与产生Ab1B细胞表面BCR相同的独特型,当Ab1产生到一定水平时,可刺激体内识别Ab1上Id的另一组B细胞克隆产生Ab2;以类似的方式Ab2刺激机体产生Ab3。由于适量抗原刺激,反应只能使Ab3产生一定水平,此水平足以抑制Ab2,但尚不能刺激Ab4。这样,使Ab1克隆细胞得以活化增殖,产生较高水平的针对入侵抗原的抗体(Ab1)。
2.低剂量耐受性 低剂量抗原刺激机体产生较低水平的Ab1,只能微弱地刺激产生Ab2,其浓度恰好能够抑制Ab1但不足刺激Ab3,因此发生低剂量免疫耐受。
3.高剂量耐受性 高剂量抗原的刺激信号强,可产生较高水平的Ab3,进而刺激产生Ab4,Ab4对Ab3的反馈抑制,导致Ab2逃脱Ab3的抑制而增殖,从而抑制了Ab1克隆表现为高剂量耐受性。
Id除分布于Ig分子或BCR V区外,还存在于T细胞受体(TCR)α和β链,包括t h、Ts、TDTH和Tc等不同T细胞亚群。TCR上的Id可用抗IdAb或抗IdT细胞来鉴定。一般来说,TCR上的Id与Ig或BCR上的Id有所差别,这可能是TCRα、β链的V基因与Ig重链、轻链V基因同源性较低的原因。抗TCr Id也能模拟抗原而活化T细胞。最近研究表明,抗原特异性B细胞上Id不仅可被Ab2B细胞所识别,也可被独特型特异性Th、Ts或Tc所识别;同样抗原特异性T细胞上Id可被TCR特异性T细胞和Ab2 B细胞所识别,这可能与淋巴细胞V基因库中,TCRα、β链V基因库与BCR免疫球蛋白重量链和轻链V基因库有一定程度的同源性有关。
图9-8 TCR和BCR上Id相互被识别的示意图