第七节 经B淋巴细胞抗原受体介导的信号转导分子基础
B淋巴细胞是另一群重要的免疫活性细胞,它有两个基本的功能:一方面作为免疫效应细胞直接参与免疫应答,介导体液免疫;另一方面作为特异性的抗原提呈细胞选择性地捕获抗原并提呈给T细胞,协同和调节T细胞免疫应答。B细胞以上的两个基本功能是通过其表面的抗原受体所介导。B细胞抗原受体的信号介导由许多分子参与,主要包括B细胞抗原受人本(b cell receptor,BCR)和BCR相关联的分子。通过信号蛋白激活B细胞内的多种酶活化途径,最终导致B细胞的增殖、活化,合成和分泌免疫球蛋白。
一、BCR的组成及功能
同 TCR/CD3复合体一样,BCR也是一种由异源寡聚体形成的复合体。目前证实BCR至少同二部分组成:(1)膜表面免疫球蛋白(surface membrane immunoglobulin,mIg);(2)Igα和Igβ(图8-14)。
1.mIg mIg是由二和要重链和二条轻链通过二硫键相连形成的四聚体结构,同分泌形式的Ig不同,mIg的重链是穿膜的多肽链。重链的胞膜外区由1个V区、3-4个C区组成,其胞浆尾部由数目不等氨基酸组成,μ和δ含3个相同的氨基酸残基,α含14个氨基酸残基,γ和ε含28个氨基酸残基。成熟的B细胞含有mIgD和mIgM两类mIg;而未成熟的B细胞仅含mIgM。mIg主要的功能是识别外源性抗原。
2.Igα和Igβ Igα和Igβ(分别为CD79a和CD79b)都是免疫球蛋白超家族结构相关基因的表达产物,又分别称为MB-1(为mb-1基因表达产物)和B29(为B29基因表达产物)。Igα和Igβ在物种之间有高度的保守性。人和小鼠Igα、Igβ在DNA水平上有90%的同源性。在人B细胞中Igα和Igβ的分子量分别为47kDa和37kDa;在小鼠B细胞中Igα和Igβ的分子量分别为34kDa和39kDa。Igα和Igβ都属于I型跨膜糖蛋白,在B细胞中以二硫键相连形成异源二聚体,并同mIg相连接。Igα和Igβ胞膜外结构域同TCR/CD3复合体中CD3的γ、δ、ε链相似,各含有1个免疫球蛋白样结构域,此外,MB-1和B29同CD3的γ、δ、ε和ζ亚单位一样,胞浆内也含有一个含酪氨酸的ARAM。Igα和Igβ的功能有二个方面:(1)作为一个主要的信号传导分子传递外界抗原结合受体所产生的刺激信号;(2)参与mIg的表达及其转运。B29(Igβ)单独足以转运IgM到细胞膜上,且参与由免疫球蛋白所介导的抗原特异性信号传导。在垂体细胞系中,Igα和Igβ可以完全导致mIg的产生,并部分重建由IgM所介导的信号转导。
二、BCR相关联的分子
1.BCR与激酶相连 除了BCR/Igα/Igβ外,B细胞表面还存在着一些BCR相关联分子,参与BCR的信号转导。BCR交联后可引起Igα和Igβ的酪氨酸磷酸化,Igα和Igβ胞浆内功能区无激酶同源序列,但Igα和Igβ确与一些激酶相关联。尽管Igα和Igβ有着高度的同源性,但它们胞浆尾部连接激酶分子的种类有所不同。Igα胞浆内同src家族激酶p56lyn和p59fyn相连结,而Igβ胞浆尾部则同磷脂酰肌醇-3激酶(PI3K)及尚末明确的磷酸化蛋白pp40和pp42相连结。
2.BCR与CD19、CD21相连 成熟人B细胞mIg可以同CD19和CD21跨膜蛋白相连。CD19是一种B细胞特异的抗原,为Ig超家族成员,表达于除浆细胞外所有B细胞上。CD21为C3dg和iC3b的受体(CD2)。抗原一方面可以与BCR结合,另一方面可以通过C3dg与CD21相连,构成了B细胞的双重抗原识别(dual antigen recognition)(图8-15)。这种双重抗原识别可以使BCR与CD19、CD21形成多聚化,为B细胞的活化提供了最佳刺激信号。此外,MHC-Ⅱ类分子,Fcγr Ⅱ以及胞浆p21ras蛋白与mIg发生共帽(co-capping)现象,从而参与B细胞的信号转导,但它们的调节作用不同。MHC-Ⅱ类分子在人B细胞转导激活信号,在LPS激活的B细胞中介导负信号;Fcγr Ⅱ的交联可以加速由mIg交联所致升高Ca2+的清除,并阻止mIg介导的信号转导给G蛋白;p21ras是一种GTP酶,可将浆膜上的生长信号同胞质中c-raf和c-erk激酶连接起来。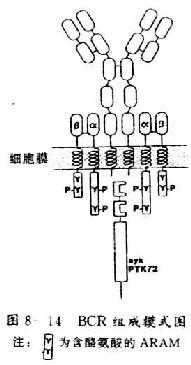
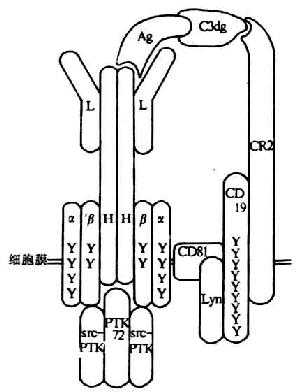
图8-15 B细胞双重抗原识别模式图
三、PCR介导的信号转导途径
BCR的交联可以激活多种酪氨酸激酶,引起许多蛋白酪氨酸磷酸化,酪氨酸激酶底物有Igα和Igβ链、LC的γ1和γ2异型、p21ras蛋白以及PI-3K等。这些分子发生要酪氨酸磷酸化后可被激活,从而介导信号的传导。B细胞活化过程中涉及到多种磷脂酰肌醇的产生及其调节(图8-16)。其中某些磷脂酰肌醇产物主要参与信号传递,如IP3为一种第二信使,引起胞浆内Ca2+浓度升高,激活Ca2+浓度升高,激活Ca2+依赖的蛋白激酶。3,4-二磷酸磷脂酰肌醇[phosphatidylinositol3,4-bisphosphate,PtdIns(3,4)P2]及3,4,5-三磷酸磷脂酰肌醇[PtdIns(3,4,5)P3]均可以充当第二信使,在体外可激活PKCζ异型。
BCR交联后通过PI-3K和PLC二种酶介导的不同的信号转导途径调节磷脂酰肌醇的产生,产生不同种的磷脂酰肌醇可分别激活下游的信号蛋白,使得信号逐级传递,最终引起B细胞的活化、增殖,并发挥其生物学功能。
图8-16 不同磷酸化磷脂酰肌醇的产生及调节
注:PtdIns:磷脂酰肌醇
PtdIns3P:磷脂酰肌醇3磷酸
PtdIns4P:磷脂酰肌醇4磷酸
PtdIns(3,4)P2:3,4-二磷酸磷脂酰肌醇
PI-3K:磷脂酰肌醇3激酶
PI-4K:磷脂酰肌醇4激酶
PtdIns(3,4,5)P3:3,4,5-三磷酸磷脂酰肌醇
Ins(1,4,5)P3:1,4,5-三磷酸肌醇
PLC:磷脂酶C
DAG:二酰基甘油
一、BCR的组成及功能
同 TCR/CD3复合体一样,BCR也是一种由异源寡聚体形成的复合体。目前证实BCR至少同二部分组成:(1)膜表面免疫球蛋白(surface membrane immunoglobulin,mIg);(2)Igα和Igβ(图8-14)。
1.mIg mIg是由二和要重链和二条轻链通过二硫键相连形成的四聚体结构,同分泌形式的Ig不同,mIg的重链是穿膜的多肽链。重链的胞膜外区由1个V区、3-4个C区组成,其胞浆尾部由数目不等氨基酸组成,μ和δ含3个相同的氨基酸残基,α含14个氨基酸残基,γ和ε含28个氨基酸残基。成熟的B细胞含有mIgD和mIgM两类mIg;而未成熟的B细胞仅含mIgM。mIg主要的功能是识别外源性抗原。
2.Igα和Igβ Igα和Igβ(分别为CD79a和CD79b)都是免疫球蛋白超家族结构相关基因的表达产物,又分别称为MB-1(为mb-1基因表达产物)和B29(为B29基因表达产物)。Igα和Igβ在物种之间有高度的保守性。人和小鼠Igα、Igβ在DNA水平上有90%的同源性。在人B细胞中Igα和Igβ的分子量分别为47kDa和37kDa;在小鼠B细胞中Igα和Igβ的分子量分别为34kDa和39kDa。Igα和Igβ都属于I型跨膜糖蛋白,在B细胞中以二硫键相连形成异源二聚体,并同mIg相连接。Igα和Igβ胞膜外结构域同TCR/CD3复合体中CD3的γ、δ、ε链相似,各含有1个免疫球蛋白样结构域,此外,MB-1和B29同CD3的γ、δ、ε和ζ亚单位一样,胞浆内也含有一个含酪氨酸的ARAM。Igα和Igβ的功能有二个方面:(1)作为一个主要的信号传导分子传递外界抗原结合受体所产生的刺激信号;(2)参与mIg的表达及其转运。B29(Igβ)单独足以转运IgM到细胞膜上,且参与由免疫球蛋白所介导的抗原特异性信号传导。在垂体细胞系中,Igα和Igβ可以完全导致mIg的产生,并部分重建由IgM所介导的信号转导。
二、BCR相关联的分子
1.BCR与激酶相连 除了BCR/Igα/Igβ外,B细胞表面还存在着一些BCR相关联分子,参与BCR的信号转导。BCR交联后可引起Igα和Igβ的酪氨酸磷酸化,Igα和Igβ胞浆内功能区无激酶同源序列,但Igα和Igβ确与一些激酶相关联。尽管Igα和Igβ有着高度的同源性,但它们胞浆尾部连接激酶分子的种类有所不同。Igα胞浆内同src家族激酶p56lyn和p59fyn相连结,而Igβ胞浆尾部则同磷脂酰肌醇-3激酶(PI3K)及尚末明确的磷酸化蛋白pp40和pp42相连结。
2.BCR与CD19、CD21相连 成熟人B细胞mIg可以同CD19和CD21跨膜蛋白相连。CD19是一种B细胞特异的抗原,为Ig超家族成员,表达于除浆细胞外所有B细胞上。CD21为C3dg和iC3b的受体(CD2)。抗原一方面可以与BCR结合,另一方面可以通过C3dg与CD21相连,构成了B细胞的双重抗原识别(dual antigen recognition)(图8-15)。这种双重抗原识别可以使BCR与CD19、CD21形成多聚化,为B细胞的活化提供了最佳刺激信号。此外,MHC-Ⅱ类分子,Fcγr Ⅱ以及胞浆p21ras蛋白与mIg发生共帽(co-capping)现象,从而参与B细胞的信号转导,但它们的调节作用不同。MHC-Ⅱ类分子在人B细胞转导激活信号,在LPS激活的B细胞中介导负信号;Fcγr Ⅱ的交联可以加速由mIg交联所致升高Ca2+的清除,并阻止mIg介导的信号转导给G蛋白;p21ras是一种GTP酶,可将浆膜上的生长信号同胞质中c-raf和c-erk激酶连接起来。
图8-15 B细胞双重抗原识别模式图
三、PCR介导的信号转导途径
BCR的交联可以激活多种酪氨酸激酶,引起许多蛋白酪氨酸磷酸化,酪氨酸激酶底物有Igα和Igβ链、LC的γ1和γ2异型、p21ras蛋白以及PI-3K等。这些分子发生要酪氨酸磷酸化后可被激活,从而介导信号的传导。B细胞活化过程中涉及到多种磷脂酰肌醇的产生及其调节(图8-16)。其中某些磷脂酰肌醇产物主要参与信号传递,如IP3为一种第二信使,引起胞浆内Ca2+浓度升高,激活Ca2+浓度升高,激活Ca2+依赖的蛋白激酶。3,4-二磷酸磷脂酰肌醇[phosphatidylinositol3,4-bisphosphate,PtdIns(3,4)P2]及3,4,5-三磷酸磷脂酰肌醇[PtdIns(3,4,5)P3]均可以充当第二信使,在体外可激活PKCζ异型。
BCR交联后通过PI-3K和PLC二种酶介导的不同的信号转导途径调节磷脂酰肌醇的产生,产生不同种的磷脂酰肌醇可分别激活下游的信号蛋白,使得信号逐级传递,最终引起B细胞的活化、增殖,并发挥其生物学功能。
图8-16 不同磷酸化磷脂酰肌醇的产生及调节
注:PtdIns:磷脂酰肌醇
PtdIns3P:磷脂酰肌醇3磷酸
PtdIns4P:磷脂酰肌醇4磷酸
PtdIns(3,4)P2:3,4-二磷酸磷脂酰肌醇
PI-3K:磷脂酰肌醇3激酶
PI-4K:磷脂酰肌醇4激酶
PtdIns(3,4,5)P3:3,4,5-三磷酸磷脂酰肌醇
Ins(1,4,5)P3:1,4,5-三磷酸肌醇
PLC:磷脂酶C
DAG:二酰基甘油