4��̼ˮ����
4.1 ̼ˮ����ķ��ࡢ�ṹ������
4.1.1 ̼ˮ����ķ���
̼ˮ����ֵ��ǡ�˫�ǡ��;��ǡ��������ࡣ�ǵĽ��������֬���ǵ��ס����������ࡣ
(1)���Dz���ˮ ��ɸ����ǵ�̼ˮ����е��ǡ�������3��7��̼ԭ�ӣ���̼ԭ�����Ķ��٣����γ�Ϊ���������졢�ѡ����ǡ����ǺͶ������м��л�����ʽ���ڣ���Ȼ�������������Ǻ����ǡ�
���Ǿ���ȩ����ͪ������ȩ���߳�ȩ�ǣ���ͪ���߳�ͪ�ǡ�
(2)˫��ÿ������ˮ��������ӵ��ǵ�̼ˮ����˫�ǡ�Ӫ�����������˫�������ǡ���ѿ�ǡ��������֡�
(3)�;���ÿ����ˮ���3��8�����ӵ��ǵ�̼ˮ����Ƶ;��ǣ�Ҳ���˰�ˮ���3��10��������20�����ӵ��ǵ�̼ˮ���������һ�ࡣ���ٷ����ࣺˮ������������Ƿ��Ӷ��������ǵij���ѿ�ǵ;��ǣ���3�������Ƿ�����ɵĽ���ѿ���ǣ��ĸ������Ƿ�����ɵĽ���ѿ���ǵȵȡ���һ��ˮ��ʱ������ֹһ�ֵ��ǣ����ӵ;��ǡ�����е��ӵ;���ˮ����������Ǻ�ľ���ǵȣ��˲��������������á��������Լ��ϳɵ��ӵ;��ǣ��к���Ҫ���������á�
(4)����ÿ���Ӹ�����ӵĵ��ǵ�̼ˮ�����Ƕ��ǡ����ٷ֦��Ͼ��ǡ���-�Ͼ��ǡ��Ӷ�������ࡣ��-�ͦ�-���Ѿ��Ƕ��������Ƿ�����ɵģ�������ķ�ʽ��ͬ��ǰ�����Ԧ�-���ռ�����ģ��������æ�-���ռ�����-�Ͼ����е��ۡ���������ԭ���ࡣ����Ӷ���ǵIJ�ֹһ�ֵ��ǣ���ʳ��ά����һ�࣬����ά���⣬��ʳ��ά����������ά�ء����������ࡢľ���ء�
(5)��֬�Ǻ�֬�ʵĽ�������֬���ɷ����ࣺ����֬������֬��
(6)�ǵ��͵����ǹ��彲��̼ˮ����͵����ʽ�Ϻ�Ļ����ﶼ���ǵ��ס��ǵ�������ܶ࣬���й㷺���������á�30����������ü�����ѧ������˲�ͬ�ķ������
1946��Stacey�ѵ����ʺ����ͣ���̼ˮ����Ļ�ѧ��ӦΪ���ij�ճ���ǣ��������ߣ���ѧ��Ӧ�Ե�����Ϊ���ij�ճ���ס�
1962��Gottschalk����̼ˮ���ϵĽṹ�ֳ������ࡣ����֮�����ظ����ֵ�˫�ǵ�λ�ĵ;���������������뵰���ʽ���߳ƶ��ǵ��ף����˰�������Protegolycans,�������һֱ�����������������ǡ���û���ظ�����˫�ǵ�λ�ĵ;��ǵ�λ�ĵ;������뵰�������ϵij��ǵ��ס�
80��������ķ��࣬�ӻ�ѧ�ṹ�������ֵú�ϸ����������ˡ����µ����ۣ���Gottschalk�ķ��෨Ϊ����
ճ���Ǻ�ճ�������֣�����Ӫ��ѧ�������г��֣���������Stacey��������в�ͬ��ճ����(Mucopolyscaccharides)ָGottschalk����ĵ������е�̼ˮ���ﲿ�֣���ư�������(Glycosaminoglycans)��ճ����ָGotrtschalk�����е��ǵ��ס���ʱ����֬���ǵ��͵�����һ��ͳ���ǽ����(Glycoconjugates)��
4.1.2 ̼ˮ����Ļ�ѧ��ɺͽṹ
(1)���ǵ����е�����Ҫ���������ǣ����Ľṹʽ�����ֱ�ʾ��������ͼ4-1��
������������̼ԭ�ӣ����������ּ�����ͼ����ͼ4-1(A)�пɿ�����2��3��4��5��̼ԭ���������ĸ���������ͬ���Dz��Գ�̼ԭ�ӡ�ÿ�����Գ�̼ԭ���������칹�壬�ǻ���������������������ͬ�Ļ����ϰ���ϰ��벮����[ͼ4-1(A)�е�6��̼ԭ��]������̼ԭ�ӵ��ǻ�������ij�Dϵ���ǡ�ͼ4-1(A)�������߿���IJ�����Dϵ�DZر��Ľṹ�����ԣ�ͼ4-1(A)��������D-�����ǡ�
ͼ4-1(A)��ʾ�Ŀ���ʽ�ṹ���Ի�������������5��ԭ�����ǻ������Ƶ���1��̼ԭ�ӵ��ǻ����ϣ�Ȼ�����ɻ�����ͼ4-1(B)����������1��̼ԭ��Ҳ��ɲ��ԳƵĵġ��ǻ�������ijƦ���D-�����ǣ��ǻ�������ijƦ£�D-�����ǡ�
��������̼ԭ���Dz���һ��ֱ���ϵģ�����̼ԭ�ӵĻ��������εġ�Ϊ�����Գ�̼ԭ�ӵĹ��ͣ��ɰ��������ʾ��ͼ4-1(C)������ƽ�洹ֱֽ�棬���߱�ʾ�ڻ�ƽ��ǰ�������ϸ�ߴ��������ڻ�ƽ��ĺ��档ͼ4-1(B)���Ҳ��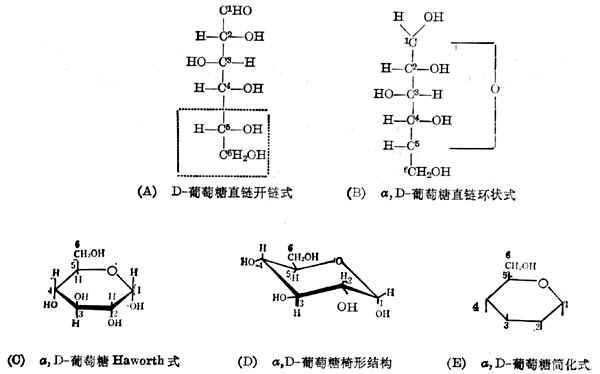
ͼ4-1 �����ǵĽṹʽ
�ǻ�д�ڻ���ƽ��֮�£������ǻ�д�ڻ���ƽ��֮�ϡ�������ʾ�Ľṹʽ����HaworthͶӰʽ��
��ʵ�ϣ���������̼ԭ�Ӳ�����һ��ƽ���ϣ�X������Ľ��˵�������������δ��ڵģ���ͼ4-1(D)���������ı�ʾ������д���ѣ�����һ�����HaworthͶӰʽ�����ҳ�����ͼ4-1(E)�������̼���ⶼʡ�ˣ�����һ�����߱�ʾ�ǻ������ڹ�������һ��ͨ�õķ����������Ǽ�������������ǵ���Glc��
��Ȼ����ڵ������ǣ������ڵ��ǡ�˫�ǡ��;��ǻ�����У�����ͼ4-1(D)����ʽ���ڡ�ֻ����ˮ��Һ�У�����Լ1%�Կ�������ʽ���ڡ�
���������ԡ���������У��е���ʹƫ����ƽ���������е�ʹ֮����������ʱ��������(+)��ʾ��������(-)��ʾ���������е�Dϵ���Ƕ��������ġ�����D-�������������ģ�����ȫ���Ǧ���D(+)�����ǡ�һ���ᵽ�������ǣ����ر�ָ�������⣬���Ǧ���D(+)�����ǡ�
������Dϵ�ġ����칹�塢�����ģ�ȫ���Ǧ£�D(-)���ǡ����Ļ������̼ԭ����ɣ���ͬһƽ���ϡ�
ͼ4-2 �£�D(-)����
�������������ǵ��칹�壬����ͬ���ǵ�4��̼ԭ���ϵ��ǻ��ڻ������棬�������ǵ��ڻ������档����Gal��
(2)˫���������ǵĻ�״�ṹ�У���һ��̼ԭ������һ���ǻ������ɺ���һ�����ǻ��Ļ�����ʧˮ����Ϊ���գ��γɵļ������ռ���
������ ��������
��������Ȼ����ڵ�һ��������Щ����Ϊҩ�
��ROH��һ�����Ƿ��ӣ�������ϵľͳ�Ϊ˫�ǡ����������Ƿ��Ӷ��������ǣ��Ԧ�-1��4�Ǹʼ�(����һ�������Ƿ��ӵĵ�1��̼ԭ���ϵ��ǻ��͵ڶ������ѷ��ӵĵ�4��̼ԭ���ϵ��ǻ���ˮ���϶��γɵļ�����д����1��4)��Ϻ�õ�������ѿ�ǡ�
�����Ǧ���D-�����Ǻͦ£�D-�����Ԧ�-1��2���ռ���ϵġ������Ǧ£�D-�������Ԧ�-1��4���ռ��ͦ���D-���������ϵġ�
(3)�;��� ��ѿ���ǵ���ѿ���ǣ����Ǧ���D-�������Ԧ�-1��4�ͦ�-1��6���ռ���ϵġ��ӵ;��ǽ��ṹ�Ƚϸ��ӣ������ǵ����м��Խ��ܡ�
(4)����
�ٵ��۵����������������Ƿ�����ɵġ���ˮ���ɺ�״�����ȵ�60��80��ɷֳ�ֱ�����ۺ�֧�����������֡�ǰ��Լռ15%������Լռ85%��
ֱ������Ҳ�������Ƿ��ӣ��Ԧ�-1��4�ռ�����һ��ֱ������������ͼ4-3��
֧������Ҳ�������Ƿ�����ɣ����Dz����Ԧ�-1��4���ռ�����ֱ�������з�֧����֧�����Ԧ�-1��6���ռ���ϵġ�ÿһ֧��20��30�������Ƿ��ӡ�ͼ4-4������ʾ��ͼ��ͼ4-5��ͼ4-4�п�����ֵķŴ����Ա�ʾ���ѷ��Ӽ�����᷽ʽ��
ͼ4-3 ֱ������ʾ��ͼ
ͼ4-4 ֧������ʾ��ͼ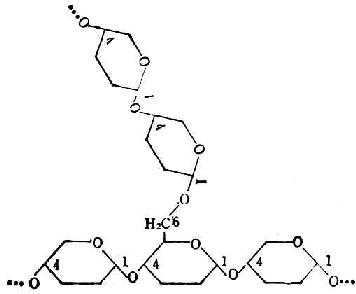
ͼ4-5 �����Ƿ�����֧�������еĽ�Ϸ�ʽ
����ԭ��ԭ��6000��3000�������Ƿ�����ɣ��ṹʽ��֧���������ƣ�������֧���ࡣÿһ��֧Լ��11��18�������Ƿ��ӡ���֧���Ǧ�1��6���ռ��������Ǧ�1��4���ռ���
�ۺ�������ѿ�;��Ƕ��߶��ǵ���ˮ����м����ṹ���Ц�1��4�ͦ�1��6���ռ���
����ά����ά���Ǧ�-�������Ԧ�1��4���ռ�����ֱ����ÿ��Լ��2800�������Ƿ��ӡ��ṹ��ͼ4-6��ͼ�������Ƿ��ӵĻ�����ǰ���������ֽ��ƽ�洹ֱ�ġ�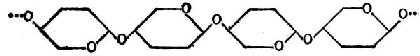
ͼ4-6 ��ά�صĽṹʽ
����ʳ��ά
����ά����D-��¶�ǡ�D-ľ�ǡ�D-�����ǡ�D-�������Ԧ�1��4�������������ģ���֧���а������ǡ������Ǻ�������ȩ�ᡣ
��������������ȩ�����ɵģ��������������ǡ��������ǡ������ǡ�ľ�ǡ�
������������ɸ�¶�ǡ�ľ�ǡ�������ȩ�����������ɣ������а����ǡ�
ľ����ľ�ʲ���̼ˮ����DZ�����ľۺ�����ں�������ʳ��άͬʱ���ڣ��������ƵĹ��ã���������Ϊ��ʳ��ά�е�һ�ࡣ
��ʳ��ά�ʹ���ά��ͬ��ֲ����ʳ���У���θ�����������������ʣ�ͳ����ʳ��ά��ʳ����������IJ������Ϊ����ά��������һ����������ʳ��ά����ʧ����õ���ֵ������ʳ��ά������20��50%��
���ǵ�������;��ǺͶ��Dz������ɵ�����ɣ������ɵ��ǵ�����������ɡ���4-1�г������ǵ����ÿ���һ����ʾ�ṹʽ�������ƺͷ��š�
��4-1 ���ǵ�������
���
�ṹʽ
����
����
������
��-D����������
��DGlen
����������
N-��������-��-D����
��DGlcNAc
��ȩ��
��-L-������ȩ��
��ltdu
������
4-����-��-D-������
��DGal-6SO3h
����һ����ɵ;��ǺͶ��ǵĵ��������������Һ�ᣬ���������������ǻ��⣬����������һ�𡣽ṹʽ��ͼ4-7������NeuAc��
�ǵ�ȩ����������������D-������(ͼ4-8)��ȩ������ԭ���ǻ����Ǵ���ɽ�洼(ͼ4-9)��
����֬��֬�Ļ����ṹ���ʰ���(ͼ4-10)�����İ�����֬����(��14��16��̼ԭ��)�������֬���ʰ���������ΪCer�����������ǻ����������������֬���͵;���(�ɼ��������30������ǻ������������)�����������֬����ȷ���ṹʽ������ʮ�֣���������������֬����IJ�ͬ��


ͼ4-7 ��Һ��
ͼ4-8 D-������
ͼ4-9 ɽ�洼
��֬���������͵����ǵ���������һ������ͬ���������ǵ��������ã��в�������֮����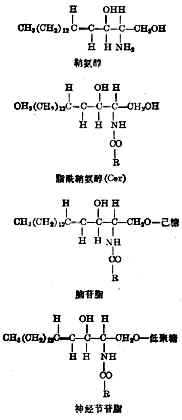
ͼ4-10��֬�Ļ�ѧ�ṹ
���ǵ����е;����ǵ����еĵ��ǻ���������������֣�
���ǣ���������(Ara)��ľ��(Xy1)��
�����ǣ�������(Fuc)��
���ǣ������ǡ������ǡ���¶��(Man)��
�����������ǣ�������������(GlcNAc)����������������(GallNAc)��
��Һ�ᡣ
��ͬ���ǵ��ף�����Щ���ǻ����������е�һ���֣���ɲ�����15������ĵ;��������ٺ͵����ʽ�϶��ɡ��Ǻ���Ϊ1��85%��
�ᵰ�����еľ۰�������һ�����Ƕ��ǡ��������н�����ֵ���ȩ��(����ȩ�����ȩ��)�Ͱ�������(���������ǻ������ǻ������������)��ϰ����˵���н�����ֵ�˫�ǵ�λ���������࣬����4-2��
���������У����������Ȼ���������������������˾۰����Ƕ��Ƕ�۵������ӡ�
�����ж��ٵ��Ƿ��Ӿۺϳɾ۰����ǣ�������Դ����ͬ����������4000��1000000���ϡ�
��4-2 �������еİ����ǵķ�������
�۰�����
˫�ǵ�λ
ȡ����
��������
��ȩ��
��������
������
��DGel
��DGleNAc
Gal;Xyl
4-����������
��DGel
��DGalNAc
GalNAc 6λ���ữ
Gal;Xyl
6-����������
��DGel
��DGalNAc
GalNAc 6λ���ữ
����Ƥ����
��DGal ��LIeU
��DGlcN
����IdU2λ���ữ ����GalNAc4��6λ���ữ��
Gal;Xyl
���������
��DGal ��LIdU
��DGlcN
GlcN���������������ữ�� ����GlcN6λ���ữ�� ����IdU2λ���ữ
����
��DGal `��LIdU
��DGlcN
GlcN���������������ữ�� ����GlcN3��6λ���ữ�� ����IdU2λ���ữ
Gal;Xyl
���������
��DGal
��DGlcNAc
����GleNAc6
GalN;Man;
Fuc;NeuAc
�����ᵥ�����ڣ�һ�㶼�ǵ�����һ�������������������У�������������⣬��ͨ��������-ľ�Ǻ͵����ʵ�˿����������ģ�һ�������Ͽ�������������
4.1.3 ̼ˮ�������������
(1)�ܽ�ȵ��ǡ�˫�ǡ��;��ǡ�����������ˮ�����۲��ܣ���ˮ���Ⱥ����ˮ���ͣ���ɺ�״�����۾��ᴦ�����ɿ��ܵ��ۡ���ԭ�ܷ�ɢ��ˮ�е����ɫ��̬����Һ������ά�ز��ܡ�����������ˮ��������ʳ��ά����ˮ���ͣ���ˮ������Դ����ΧҺ���PH������Ũ�ȵȶ���ͬ�������������5��������������ˮ����ˮ����ʳ��ά�ʺ���״��ϸ����һЩ�����ܴ����ȥ����֬�ǵ�һ����ˮ��֬�ʵ�һ����ˮ���ǵ��͵����ǵ���Ҳ����ˮ��
(2)����������ǵ������100��һЩ�Ǽ��Ǵ�����ȼ���4-3������ת���������ǵ�ˮ�����������Ǻ��Ǹ�50%��
��4-3 �ǵ����
�ǻ���������
���
����
173
ת����
130
����
100
������
74
ɽ�洼
60
��¶��
50
������
32
��ѿ��
32
����
16
4.2 ̼ˮ�����������л
4.2.1 ��������
̼ˮ����Ҫ�������Dz����ա������Ĺ��̾���ˮ��Ĺ��̡���ѿ�ǡ����ǡ����ǡ���ѿ�;��Ƕ������������������Ķ��ǽ�����һ�֣���ԭ���Ƴ�ʳƷʱ�Ѳ������ˡ�
�����ӿ�ǻ��ʼ����ǻ������Һ����ø��ˮ�⽻���1��4�Ǹʼ���������ˮ���1��6���ռ������ڵĦ�1��6���ռ������������Ǻ�������ѿ�;��Ǻ���ѿ�ǡ�
θ��û���������۵�ø����Һ����ø������PH��6.6��6.8����ʳ��û�б�θ���к���ǰ���ܳ�������һ��ʱ�䣬ʹ���ۺ͵;�����������һ���֡�
С��������Һ�Ħ�-����ø�������ú���Һ����ø��ͬ����ֱ��������������ѿ�Ǻ���ѿ���ǣ�֧��������������ѿ�ǡ���ѿ���Ǽ���4��9�������Ƿ�����ɵĶ��Ц�1��6�ռ�����ѿ�;��ǡ�
��ճĤ��Ƥϸ����������ϸ����ÿһϸ��Լ��3000����ë����ë��Ŀռ����Ч�뾶Լ0.4nm��ֻ���������������ܹ�ͨ��������ëĤ�ϵ�ø��Ӧ��Ĥ�ϵ�ø�����֣��٦�1��4����ø���������Ƿ�������������һ��������������������ѿ��ø��ˮ����ѿ�;��ǵĦ�1��6���ռ���������ø���������ǣ��ܦ�-������ø���������ǡ�
�����������������У��ٳ�ǻ�ڵ�������������˫�Ǻ���ѿ�;��ǣ�����ëĤ�ϵ������������ǵ��ǡ�
��ë˫֬�ʲ��(���)������øǶ��˫֬�ʲ��ڣ�����λ����Ĥ�⡣�������������ȫ��Ƕ��Ĥ�ڵ����䵥�ǵĵ����ʣ����������������ղ������̿��Ա����䵰������ϡ�
���䵰���ڽ����������ǰ���Ƚ�ϳ�ǻ�ڵ�Na+���볦ǻ����ǻ��Na+������ϸ���ڣ��ͷŵ������С�Na+���볦ǻ����ǻ�е�Na+Ũ�ȱ�ϸ���ڸߣ��Ե�Ũ���ŵ���Ũ��Ҫ��������������������ATP��Ӧ���ǽ���ϸ����Լ��15%���س�ǻ��25%��ɢ��Ѫ��60%�뿿����Ĥһ�˵���Ĥ�ϵ���һ���嵰��϶��뿪ϸ������һ��ϲ���Na+���������������ǵ��ٶȱ������Ǵӳ�ǻ��������ϸ�����ٶȿ죬���������Dz���������ϸ����������Ӷ����������Ч�ʡ���ʳ�ӵ���ճ��²�ʱ��95%��̼ˮ���ﶼ�������ˡ�
4.2.2 �����
С�����յ�̼ˮ������Ҫ�������ǡ����ǡ����������֣����ž����͵����ࡣ�����ǽ����ϸ���������ᷴӦ����������-6-���ᣬ����ϸ���ڵ�������Ũ�ȿ�ά���ڵ�ˮƽ��ʹ�����Dz��Ͻ����ϸ���������յ��������У�60%�����ڸ��ڴ�л���������ѭ�������ǺͰ������ڸ���ת��Ϊ�����ǡ�
�������ڸ��ھ��ֽ��л�ṩ��������Ҫ������������ĺϳ���ԭ�����ڸ��ڣ����ж����ת���֬�����͵�֬����֯����������
�����ǵķֽ��л�������Σ�
(1)�ͽ������Ƿֽ�ĵ�һ�������ɱ�ͪ�ᡣ��һϵ�з�Ӧ��̼ˮ���������⾭��ĸ���������ɱ�ͪ����ͬ�����Գ�Ϊ�ͽ⡣����ͬ���Ƿ���ʱ��ͪ���һ�����������������ڣ�����ʱ��ͪ�������ɶ�����̼��ˮ������ʱ��ԭ�����ᡣ
�ͽ��ʮ������(ͼ4-11)��Ϊ��Ҫ�������ǵ�6��̼ԭ��һ��Ϊ������ʹ���������ữ�����ܶ��г������ӽ���ȵIJ��֣�Ȼ����ȥ����ɱ�ͪ�ᡣ
������Ӧ����ϸ�����н��С�ÿһ�������ض���ø�μӡ��ڢ١�����������2����ATP��˵�������Ƿֽ��л��ʼʱҪ���������ƶ����ޡ�������������2����ATP�������ѵ���ͪ�ᣬ����2����ATP���ͽ�������������ࡣ
ϸ������NAD+��NADH�����������㶨�������е�NAD+�����NADH���ͽ�͵�ֹͣ��Ҫʹ���Ѽ�������������ͪ�ᣬ�����NADH����ת�Ƶ������������ȥ������ʱ�⾭������������ˮ������ʱ�⽻����ͪ��ʹ������⡣�������ʱ�뿪ϸ����Ѫ����������ȱ���߿ɷ�����л�����ж���
(2)��ͪ����������øa ����ʱ����ͪ����������塣�ڱ�ͪ������øϵ�İ����£��������dz�������������øϵ�İ����£��������dz�������øA��ͬʱ����1molATP��1molNADH��ά����B1B2��������������μӡ���Ӧ�Dz�����ġ�
(3)������ѭ��������øA�ϵ��������������ǵ�̼ԭ�ӣ���������ѭ������2molCO2����������ǵ���������Ӧ���������н��С���һ���̷�9��(ͼ4-12)��
������ѭ���в���3molNADH,1molFADH2��1molGTP��
NADH����ͨ��FAD��ϸ��ɫ�ؽ�������ͬʱ�ų�������ÿmolNADHͨ����һϵ�з�Ӧ���ų���������220kJ(52.6kCal)����Щ������ATP����ʽ����������ÿmolATP�ܲ���30.5Kj(7.3kCal)��������ÿmolNADHRNUD���������������������ϳ�7molATP������ָ�ڱ�״��(25�棬1������ѹ��������Ͳ���Ũ�ȶ���1M/LPH7)��˵�ġ���ģ�����ڵ�����ʱ��ʵ��֤��ֻ�ܺϳ�3molATP�����÷�Ӧʽ��ʾ��
NADH+H++1/2O2 NAD++H2O+����
NAD++H2O+����
3ADP+3H3PO4+����3ATP+3H2o
ǰ��һ����������Ӧ������һ�������ữ������ż����һ�𣬳�Ϊ�������ữ��
ͼ4-11 �ǵĽͽ�
ʵ��Ҳ֤��FADH2����ʱ����2��ATP��GTP������1molATP��������Щ��ֵ����������ȫ����ʱ�������õ������ɹ������£�
�ٽͽ⣺�����ǡ�2��ͪ��+8ATp
�ڱ�ͪ��������������øA��
2��ͪ���2������øA+2CO2+6ATp
��������ѭ����
2������øA��4CO2+24ATp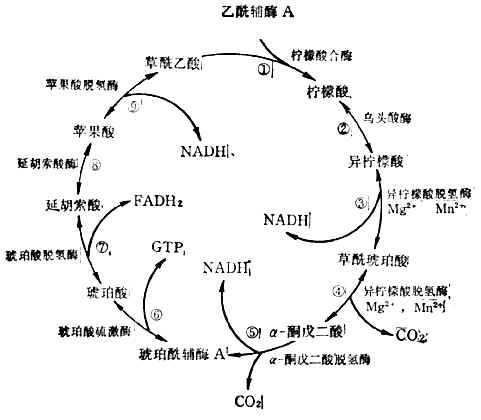
ͼ4-12������ѭ��
��ÿmol������������������6molCO2ʱ�ɲ���38molATP������������õ�������38��30��1140kJ[(38��7.3)��277kCal]��ÿmol������������6molCO2��ˮʱ�������ܵĸı���2870KJ(686kCal)�����ԣ���еЧ��=1140/2870��10=40%��
�������ñ�״�������ּ���ģ������ڸ�һЩ��Լ60%��
��������ɵ�����Ҳ�ɿ���̼ˮ�����ṩ�����������������������У���������ѭ����ࡣ֬���͵��������Ҳͨ��������ѭ��������ʳ��������ṩ��������90%����������ѭ����
�����Ǵ�л�����һ��;������ͨ�����Ƕ����ᣬ��������һ����ͨ·�����ڲ����˘y���ֽ�������-����ͨ·��
(4)����-����ͨ·��һͨ·Ŀ�����ṩ�ϳɺ�������ĺ��ǣ��Լ��ϳ�֬���ᡢ���̴��������������Ӧ��NADPH(��ԭ��ø��)(ͼ4-13)��������Լ��30%�������Ǿ�����һͨ����л����ϸ������һͨ·Ҳ�ܻ�Ծ������ʧ���ɵ�����Ѫ��ƶѪ����Ӧ����ϸ�����н��С�
����-5-����ȿ����ں��������ϳɣ��ֿ���5-����ľͪ�Ƿ�Ӧ���ɸ���ȩ3-�������-6-���ᡣ��������һ��·��Ҫ��ϸ��������NADPH�ͺ��ǵ���Ҫ������������ϸ��NADPHҪ�ö࣬��ʣ�����ǿɾ�����ȩ-3-�������-6-����ͽ⣬���ɹ���-6-����ת���������-6-�����������һͨ·��
��һͨ·��ά����B1�μӡ�Ҫʹ��·��ͨ����ʳ������������ά����B1��5-�����ͪ��ת��Ϊ������-6-����ķ�Ӧ������������ά����B1�μӡ�Ҫʹ��·��ͨ����ʳ������������ά����B1��Ӫ��״����
(5)��ԭ�����ǵ�����ϸ�����������Ժ���Ŀɴ�������������ϸ������ѹ�����ߣ�ϸ���������Ǻϳɴ���ӵ���ԭ����Ҫ����ʱ��ԭ�ٷֽ������ǡ������ԭ�����������ǣ�����ά��Ѫ�ǵĺ㶨������ԭ�����������ǣ�ֱ�����ڹ���������
ͼ4-13 ����-����ͨ·
4.2.3 ����Ӫ����л�Ĺ�ϵ
��ϰ�������ͣ������Ϳո��������������Ҫ�߱��ܴ���̼ˮ���������ȱ��ʱ��֤������Ӧ�����ױ��졣
�ͺ�Ѫ�����ظߣ��ȵ��ط������ӣ��ȸ�Ѫ���ط��ڼ��١�ʹ����������ǽ�����ࡢ�����֬����֯����ǿø�������������ѵ���������ԭ������ԭ�ĺϳɡ�������ԭ�Ĵ���������ɰ������Ǿ�������DZ�ͪ��ԭ�ɸ���-3-���ᣬ��������øA�ϳɵ�֬���ᣬ����NADPH�ṩ������������֬�����ٺ͵����ʽ�ϳɼ����ܶ�֬������Ѫ�����͵�֬����֯���档�����̼ˮ����Ҳ�ܺϳ�ijЩ�DZ��谱���ᡣ
�ͺ�Լ4h����������ʳ���е������ǡ��ȸ�Ѫ���ط������ӣ��ȵ��ؼ��٣�תΪ���ø���ԭ�Թ���Ѫ�ǡ�����ԭ���������࣬�����ܶ��õIJ���70g,����һ�����ϵ����ġ����ԣ��Ӳͺ�8h��ʼ����������֬�����ṩ���������Ϳɱ���ǣ�����֬������ռ�������ࡣ֬����ֻ�ܹ��ܣ�����ת��Ϊ�����ǡ���ˣ�Ѫ�ǵ���Դ���ò�ת�����ˡ�һ��˵��������������������⣬���������ᶼ���ܳ��ǵģ����������Ĺ۵㿴��ֻ�б�����Ȱ�����������ǵ���Դ����ʱѪ�ǵ�90%���Ա����ᣬ10%���ԹȰ��ᡣ
�����ܰѼ����������ı��������óɱ�ͪ������������ǡ��ո�ʱ�������н���ѪҺ�İ��������ӣ���������30��40%�DZ����ᡣ�����⵰���ʵı����Ậ������10%������ı�������������ģ������Ǿ��ͽ������ͪ�ᣬ�پ�ת�����ñ����ᡣ�����������ǵ�����û�����ӡ����ӵı�����ʵ���������������ᡣ
����ѹȰ���ȥ����æ�ͪ����ᣬ�ٺ������ij��ǰ������������ת�����ö��ñ�ͪ�ᣬ���������ǡ���-ͪ�����Ҳ�ܾ�������ѭ���γ�ƻ���ᣬ���������ǡ�
4.3 ̼ˮ�������������
4.3.1 ��������
(1)������ÿmol��������������ʱ������������2870Kj(686 kcal)�������ǵķ�������180��ÿ�������Dz���2870/180=15.94kJ(686/180��3.81kcal)����ͳ��ȡ������16Kj(4kcal)������������֯���������Ƕ�Ϊ16KJ(3.75kcal)������Ϊ18Kj(4.1kCal)���ҹ�������ҩ��涨������Һ�õ������Ǻ�1mol�ᾧˮ��ÿ�˲���16��180/198��14.5kJ(3��75��180/198��3.41kCal)��������Һʱ�������ṩ������Ӧ��14.5Kj/g(3.41kCal/g)���㡣
(2)��ʳ��̼ˮ������ռ�����İٷֱ�̼ˮ����û�й涨����Ҫ������Ϊ��������������ڶ��ܱ���ǡ���ÿ��ÿ����������ʳ����������̼ˮ����Ϊ50��100g�����������ͪ������֯���ֽ���࣬�Լ������ӣ��ر��������ӣ���ˮ�Ķ�ʧ��
ʳ����̼ˮ����̫��Ҳ���á�����Ӫ��ѧ�ҵ�������������е�55%Ӧ��̼ˮ��������Ӧ�����е���˫���ṩ�IJ�����14%��
�������(��Ҫ�ǵ���)��ͬʱ���ܻ�õ����ʡ�֬�ࡢά���ء������ʡ���ʳ��ά�����뵥˫��(��Ҫ������)ʱ������ȡ�ó������������Ӫ���ء������������ǹ���������ȣ�ݡ���Ѫ�ܼ���������
(3)���ǹ����Ӱ��
�ٹ��IJ������ǹ��࣬��ʳ��ά���٣����IJ��������ʸߡ��籾���������������˵�������Խ��Խ�ͣ������IJ�������ȴ�������ߡ���1900���ÿ10������167.3�ˣ�����1965���237��7�ˡ�
��1909�굽1965�꣬�����˵�ʳ���У�ÿ��ÿ����ȡ�������½�Լ10%�������ʵı仯����֬������������ռ�İٷ�����������9%�������ӵ���Ҫ��������֬����(��������Ϊ����)����ʳ���м������õĵ��̴���������1909��1913��ÿ��ÿ����495mg��1965����518mg������֬���͵��̴��仯���������Եı仯�ǵ���������٣�������˫�dzԵö��ˡ�1909�ꡫ1913��ÿ��ÿ����ȡ̼ˮ����492g����������ռ31.7%��1965��̼ˮ����374g������ռ51.2%��
�ٴ���ʳ��ά�ĽǶȿ���������������������˵�������Ҳ�������١����dzԵ�ˮ�����߲˵��������䣬ˮ����Ʒ�ִ�ƻ����Ϊ��Ը̽ۡ�ƻ���������࣬�̽۵Ĺ��������ڽ�Ƥ�У�һ�㲻�ԡ���һ�ı䵼������ʳ��ά�������ļ��١�
�Ӻ�ķ��棬Yudkin�Ƚ���15�����ҵ���������֬��������֬����ֲ��֬�������͡��������͡��ܵ��ס����ﵰ�ס����ǵ���ȡ�����IJ������ʵĹ�ϵ������ֻ�����dz�����ء�
�ݺ���������ıȽϣ���˵����ȡ�����IJ������ʸߣ���ʳ��άҲ��һ��Ӱ�졣����ʵ���о�Ҳ��֤����һ�㡣
������Cohen�����ƾ���ɫ�кͳ��ڶ�������ɫ�е�Ҳ���˵��������ʺ�̼ˮ�������������˱Ƚ�(��4-4)��˵������������ߣ������ʸߵö࣬��̼ˮ�����������������������г��벻��
��4-4 ������̫��̼ˮ�����������������ʵıȽ�
���
������(kCal)
̼ˮ��������
����
�������ʣ�%
Kcal
�ƾ�10��������
2237
343
1372
6.6
26.4
0.06
�ƾ�25��������
2559
377
1508
63.0
252.0
2.90
���Ƕ�Բ��ã���ô����Ӧ�Զ��٣��Խ����ˣ�ÿ��ÿ��������2g�������ڲ�������Ѫ�ǡ�����������֬���ᡣ���ಡ�������ߣ��������ṩ������ռ��������5%��������
(4)Ӫ������ʱ̼ˮ�����ѡ��ڷ�̼ˮ���ﲹ������ʱ������ѡ����˫�ǡ����蹩��8400Kj(2000kCal)����������ʱ��586g�����˶�ˮ����������3L����˱�ȻҪ�ý�20%����Һ����������ѹ��Ѫ���4�����������͡���ʹ����к����ѡƽ����������������5������ѿ�;��ǣ���25%ʱ����������������������Ը����á����ֵ;��Ǻ������������ա�
��Χ������Һʱ����ѹ���ܸ���500mOsm��kg-1ˮ�����������Ѫ˨�Ծ����ס�10%������(��ѹ500mOsm��kg-1ˮ,)ֻ�ܶ�ʱ��ʹ�á��Ǵ�����ѹ�ϵͣ�������ע���ˮ�洼����20��40%���������ų�������10%ɽ�洼����Χ����Ѫ˨�γɵĿ����ԣ���10%�����Dz�ࡣľ�Ǵ������������Ͳ������������ڵij����������°��ı�����Ҳ��������ѿ�;��ǣ���Ϊ��ͨ�����࣬�;��Dz���ˮ��������ǹ�������֯���á������辭��Χ�����ṩ��������ʱ��ֻ��Ӧ��֬�����������Ʒ��Intralipid����û�ж��ٸ����ã��ѹ㷺�����ٴ���
���ľ�������Һ��ܿ��ܱ�ѪҺ���ͣ����ؿ�����ѹ�����⣬���øߴ�25%��50%������Ũ��Һ��
4.3.2 ����ϸ������֯
ÿ��ϸ������̼ˮ����京��ԼΪ2��10%����Ҫ����֬���ǵ��͵����ǵ���ʽ���ڡ��ֲ���ϸ��Ĥ��ϸ����Ĥ��ϸ�������Լ�ϸ��������С�
ϸ��Ĥ�����㡣����DZ�Ĥ����Ҫ���ڲ��������������ɡ��ڲ���Ĥ��������֬��(��֬����֬)���ж��ɡ�֬�ʵ�֬�����ǷǼ��Եģ��������ڣ��ڲ�����⣬����֬��˫��(ͼ4-14)���ǡ����ἰ�������Ի��Ŷ�����Ĥ�����ࡣ��Щ������Ƕ��֬��˫���ڣ���ϵ�ǵ����ǣ������ἰ�������Ի��Ŷ�����Ĥ�����ࡣ��Щ������Ƕ��֬��˫���ڣ���������Ĥ�ϡ�Ƕ��Ĥ�ڵ��г���άϸ��������ϸ����Ƥϸ���ϵ���������أ�������Ĥ�ϵ��и�ϸ��������������������������ء��������£�֬��˫����Һ��ġ�֬�ʷ��Ӻ͵����Ӷ�������Ĥ���ƶ�����Ĥ����ֵĺ�����ϸ������ͬ����ϸ����Ĥ��������65%��֬��30%��̼ˮ�����5%��
��ÿ��ϸ������̼ˮ�����⣬�ǽ���ﻹ�㷺�����ڸ���֯�С��Ժ�����֯�к�������֬�����ݷֲ��������ϡ������١�θ��Ƣ���Ρ��Ρ����١�����Ĥ����ϸ������ϸ���ȶ�����֬��
�����������������ڵ�ճҺ�����ǵ��ס��Ǻ����е���ճ���ף�Ѫ���е�ǰ���ס���1-����2-����-����-�ס���Ѫøԭ����ά����ԭ���������ף������еļ�״���ء��ټ�״�ټ��ء������ټ��ء��ٺ�ϸ�������أ�ø�еĵ���ø������ø������ø��ˮ��ø�ȶ����ǵ��ס�
������������ڹǡ����ǡ����졢�ʹ�����Ĥ��Ƥ����Ѫ�ܡ�������ؽ�Һ������Һ�С�
�����֯��ϸ������ʣ���Ҫ�ǽ�ԭ�͵���������ɡ�
4.3.3 ������Ϣ
����������һֱ��Ϊ������Ϣ�ǵ����ʺͺ������,̼ˮ������Ҫ�����ǹ������ܺ�����֯����30�����������˴���ʵ��֤�ݣ���1968���ѹ���̼ˮ������Я����Ϣ���Ժ����кܴ�չ����������ڣ�
ͼ4-14 ��Ĥ���ɵ�ʾ��ͼ
��֬��˫�㣬�ڵ�������Ի������ߴ���֬������
bǶ��Ĥ�ڵ���֬������������Ĥ�⡣ cǶ��Ĥ�ڵ��ǵ��ף�����������Ĥ��
d������Ĥ�ϵĵ����ǡ� e���̴� f�����ʷ���
(1)��Ϊ�ǵ���ϸ����ʶ��ı��ϸ����ϸ�����ܹ��ʶ��ġ����ǿ��ϳ��Է��Ǻ��Լ���ͬ�ģ���������һ�����ģ�������������һ����ġ���Ѹ��ࡢ�������̥����ϸ������ͬһ������������ǻ���Լ�����һ����Ϊ�������ϳ�����ڶԷ�ϸ��Ĥ�ϵ������Ǻ��Լ���ͬ�ġ����罫һ���˵�������ֲ����һ��������ȥ�����ܵ������߰�ϸ�����ų���ƻ���ԭ��֮һ���������ϳ�ֲ������ϸ��Ĥ�ϵ��������������IJ�ͬ��
�ڽϼ������У��ɰ��ǵ�ʶ�����ÿ��ø������
��������Щ�ǵ��������һ�������Ҫ���������ϸ���ϻ���ҲҪ�����������ڷ������õ��ǵ���ϸ�������������ڡ����ߵ����������������ϻ������������Ľṹ���˸ı䡣��ϸ����Ĥ�ϵ�������ʶ�����ָı��˽ṹ��������������ǣ�ʹ���ǽ����ϸ���ڣ�����ø����ʹ֮���⡣���ڷ������õĸ�ϸ������ϣ�����������١�
���鶯������࣬������������ʶ���ǵ������壺
������Һ�����ǵ����������˵�������GΪ������;�������ĩ������Һ�ᡣ��Һ�ᱻø���Ժ�ĩ���ǰ����ǡ���ϸ����ʶ���ĩ���ǰ����ǵ�������G���������ĩ������Һ���������G��
��������������Ѫ���ǵ�����֪��21�֣�����ϸ����Ӧ�ĵ����ߴٺ�ϸ�������ء������ݳ��켤�ء���ëĤ�����ټ��ء������أ����嵰����Ѫ��ͭ�����ף�Ѫ���ؽ�ϵ��ס�ά����B12���ݵ��ס�Ƥ������ס�Ѫ���ǵ��ס��������ǵ��ס���Ѫøԭ�ȡ�
������屾��Ҳ���ǵ��ף�������41000������10%��
�ϻ��ĺ�ϸ��Ĥ�ϵ�����Һ�������������ĺ�ϸ��Ϊ�٣��ܱ���ϸ��ʶ��������ɡ������ĺ�ϸ����������ĩ������Һ�ᣬ�����ϣ����������ڷ������á�
�������������ڸ�ϸ��Ĥ�ϣ���������������ס�
�������¶�������ڸ�ϸ��Ĥ�ϣ���������������ס�
�ܸ�¶�ǻ�N-�������������������ڿݷ�ϸ�����档
��Щ���廹������غ�衢Ƣ�ࡢ�κ����С�������ֹ�ĺ�ϸ��Ҳ�ܱ�Ƣ��ʶ�𡢽�Ϻͽ��⡣
̼ˮ����������������ʶ�����ã����������ڱ����ڻ������ȶ������һ���Ӧ�����ٴ����磺A�����ҩ������������Եķ��ӣ������������Ϊĩ�˵ĵ;������ϣ�������ѡ���Եص���ijЩϸ�������ϸ���ϣ�B��ϣ�ȥ����Һ����Ѫ���ǵ��ף��������ر��ⶨ�����������۸�Ӳ�������ס�ԭ���Ըΰ��и�ϸ�������˳̶ȡ����������ĺ�����Һ����������ı�ֵ�����������Ƹ������Ĵ�С��
(2)�п�ԭ������Щ�;����п�ԭ���á��������е�����;�����Ϊ����Ķ��Ľ�Ϻ�ע�������ڣ��ܲ������塣���ֿ�����ʶ����Ϊ��ԭ�ĵ;��ǡ������ṹ���иı䣬����Ͳ�����������DZ������п�ԭ���á�
����Ѫ�ͷ�����Ǻ�ϸ��Ĥ���ǵ�����֬�ĵ;�����ĩ�˵��Ƿ��ӡ�����������14�����Ƿ�����ɣ�ĩ��4����ͼ4-15��A��Ѫ����ĩһ��������N-�������������ǣ�B��Ѫ���ǰ����ǣ�AB��Ѫ�������ֿ�ԭ�����У�O��Ѫ��û�����������Ƿ��ӣ�����ֻ��13�����Ƿ��ӡ�
ͼ4-15��ͬѪ�Ϳ�ԭ�ĵ;�������ĩ���ĸ�����
����ϸ����Ҳ������ԭ���õ���������NILϸ����һ������������ĩ������Һ�ᡣ�����������ϸ�����Ի��Ժ�����ĩ��ȱ���������Һ����ӡ��Ѷ��Ի���NILϸ��������ע����ڷ���������ͬʱ������Ѫ���в�������ȱ��Һ�����֬���������Ͳ����Ϸ�Ӧ�Ŀ��塣���Ŵ���Ĥ�Ϸ�����֬���ǵ����ֶ�Խ��Խ��ϸ���Լ���������ѧ��ҽѧ�Ľ��������ԭ����ķ�Ӧ�����п������ڰ�֢�����ơ�
(3)��ϸ����ϸ����ճ���з�������ϸ����ϸ����ճ����һ��ճ�ŵķ�ʽ�����֣�
��ֱ��ճ��ϸ���������ǻ�ת��ø����֬���ǵ��ף����ø������(�������)��ϣ�ͬʱ���Ǽ�����һϸ������֬���ǵ����ϣ�ʹ������ϸ��ճ����һ����ͼ4-16������ϸ����ø����֬���ǵ��ľ���Զ�������ڶ���һ��ϸ���Ͻ�ϣ�ֻ������һ��ϸ�����ϡ�����ϸ�����߽Ͻ�����������ϣ���������ϸ������ת�ơ�������ϸ����ѪС�塢�����Ͷ��Ի��ij���άϸ����֤ʵ����������ǻ�ת��ø����-��������ø���ƻ�����ճ�����ã�˵��ͼ4-16�е��ǣ������ǰ����ǡ�
��ͨ��ϸ��������е�һ�����ʣ�������ϸ������ѷ���õ�����CSP�ǵ��ף�LETS�ǵ��ס������ǵ��צ�����ճ�����ȡ�����ϸ���ܲ���CSP��LETS�����Ի�����������ϸ��ճ���١����ڶ��Ի��ļ��߳ɹ�ϸ��������ϸ���������м���CSP��LETS�����������ǵ�������
ͼ4-16 ϸ��ճ��ʾ���ͼ
e ����ø G �����ǵ�����֬ S ������
ϸ��ճ��ʧ���ɴ����������˵�ѪС�������һ���ǵ��ף��;�����ĩ������Һ�ᡣѪ�����Ѻ�ѪС���ͨ����Һ��ճ���ڱ�¶������Ѫ����Ƥ�ϣ���ֹѪ���á�BernardSoulier�ۺ�������ѪС��Ĥ�������ǵ������ܵͣ�����ֹѪ��
(4)ϸ���ĽӴ�����ϸ������ʱ������ϸ�����ӵ�һ�����ܶȾͲ������������ֶ��������������ã��нӴ����ơ�����ϸ��ʧȥ�ĽӴ����ƣ�����������ܶȱ�����ϸ���ߵöࡣ�������ƺ�ϸ������������кܴ�Ĺ�ϵ��
ϸ��Ĥ�Ļ�ѧ�ṹ���������ʹ�л�IJ�ͬ��������ͬ����ϸ������ʱ��һЩ�����DZ�¶�ģ�������ϸ�������ڵ���̥ϸ��������ϸ������˿����ʱ��ijЩ����Ҳ�DZ�¶�ģ�������˿���Ѻ�Ķ�ʱ���ڣ��ܺϳ�һ�㵰���ʰ���Щ�����ڸ�������ʹϸ������Ӵ�����Ĥ�ϵ������ỷ��ø�������ӣ�ϸ����cAMPŨ�����ߣ�����Ϣ����ϸ�����ϣ�ֹͣDNA�ĺϳɡ�����ϸ���ܺϳ���㵰���ʣ����ߺϳɱ߱�����øˮ�⣬���Բ�����Ч�ؽӴ����ƶ��������á��ȵ���ø��ˮ������ڸ��õĵ����ʣ�ʹ�����ij���άϸ��ʧȥ�Ӵ����ƶ������������˱��ʰ�����Ƥϸ��������ϸ������ϸ�������ܶ�����cAMP(cAMP��һ���������cAMP������ϸ��)�����ƣ�������һѧ˵��֤�ݡ��������⣬ϸ�����������ṹ�ĸı�Ҳ��ʧȥ�Ӵ����Ƶ�һ��ԭ����֪��������ϸ�����ǵ�����Һ�Ậ�������ӣ�����֬�ĺ������ٵȡ�
4.3.4 ��������
�ǵ��͵������л������á��ؽ�Һ���д��������ᣬ�ǹؽڻ����������Һ�е��ǵ��ף�ʹʳ�������ƶ����ҿɰ���ʳ�Ӻͷ�㣬ʹ��ճĤ���ܻ�е�ͻ�ѧ�IJ��ˡ����������ǵ��ף��з�ֹ֧���ܺͷ�����Ƥ����������������������������������á���ֳϵͳ���ǵ��͵����ǣ��������ھ����˶���������̥�����á�
4.3.5 ���������ʲ�������ø����
��Щ�����ʣ���ø������Һ�е��ǵ��ף�ƽ����������ø�������������ڷ����е������ڱ������ǡ������ṹ�ĸı䣬���M���ٷ��ڵ��ǵ���ȥ��ĩ�˵���Һ������������ĵ���ø������
4.3.6 ����ϸ��Ĥ��ͨ��
�������Ĥ����������������Ի��ţ��ܿ���ˮ���ӡ������Ӻ�С���ӵ��л�����ƶ��ͽ���ϸ���ڲ������跨ʹ�������ܺϳɵ�������ȥ�������ǾͲ��ܽ��뼦�߳���άϸ����ȥ���Ǵ�л�����ϰ���
4.3.7 ���ʽ�ʡ����
ʳ������̼ˮ���ﲻ�㣬���岻�ò��ӵ�����ȡ����������Ϊ������Ҫ�������г�������Ӫ���ء���Ҫ����ȵذѰ��������ڵ����ʺϳɣ�����ȡ���谱�����ͬʱ��һ��Ҫ���㹻��̼ˮ���﹩Ӧ��
4.3.8 ��֤֬���ij������
ʳ����̼ˮ���ﲻ�㣬����Ҫ���ô����֬���������������������֬���������������һ���ȡ�����֬�����࣬��ֽ��л���м����(ͪ��)������ȫ�������������ͪ������ʳ�е�̼ˮ����ɱ�֤����������������
4.3.9 ��ⶾ����
���е�������ȩ���ܽ��һЩ�����Ļ�����Լ�ϸ�������Ķ��صȣ���ͬ�ų����⣬�ⶾ���á�
4.3.1 0��Ϊ�ϳ��������ӵ�ǰ��
������������������̼ˮ�������ϳɵģ������ʡ���ड�ijЩ�����ᡢ߲�������̴��ȡ��������Ǻϳɺ��ᡢ�����ʼ����̴���һЩ�����ʳ���е����������õú��١���ɺ���ĺ��Ǻ��������������������кϳɵġ�
4.3.1 1��ʳ��ά����������
�˲���������ʳ��ά�����᳦��ϸ����ø��ʹ��ά�ء�����ά�غ����ֽ⡣���Դ�����ų�����ά��ֻ��ʳ���е�20��70%������ά��15��45%������Լ10%������Լ70%���ֽ�������⡢������̼�����ᡢ���ᡢ����Ͷ���ȡ��ֽ�ʱҲ���������������������е�һ���֡�����������������ġ�
���䲻��������ʳ��ά������������һ�����������á��ڿ�ǻ����Ӿ����̼���Һ�ķ��ڡ���ʱ���ӳ���Ҳ����θҺ�ķ��ڡ���С��������Ҳ��Ӱ�졣����൱��1004kJ(24kCal)��ƻ����ƻ��֭����һСʱ��δ�ָ�����Ϊƻ��֭�е������պܿ죬�̼����ȵ��صķ��ڣ�ƻ�����й�����ʹ�������գ������˹�����ȵ��ط��ڡ���ʳ��ά�������ӵ�֭�ķ��ڣ�������֭�����ʳ�����ͨ����ʱ�䣬���ӷ�����������ٷ��Ӳ�ȡ�
4.4 ��ʳ��ά��һЩ�����Ĺ�ϵ
�����˲���������������ʳ��ά�����Գ���������Ϊ���dz����ӳ��䶯����ֹ�����⣬�������á�1956��Cleave�����������е�ijЩ��������ʳ���г�ȥ����ʳ��ά(�Ծ�����ۺ�������̼ˮ�������Դ)��������ʮ������ڷ�չ�й��ҹ���������ҽ��������ijЩ�����ڵ��ؾ�����Զ�Ϸ������Ϊ�١�1972��Painter��ʳ�������������������Ҳ��õ����õ�Ч�����������˶���ʳ��ά�����ӡ�
4.4.1 ����ʳ��ά�йصļ����У�
ӡ�ȳ��о����Ծ�����Ϊ����ũ���Dz��ס��ͻ�˹̹�����˳Ծ����棬ũ����ȫ����Ʒ��������������������dz��и���ũ�塣
С���������д���άռ5%ʱ���ָ�Ѫ�ǣ���������>75%ʱ��
�ô���ά��������������в��ٵı��������磬13����ҩ���Ƶ������ˣ�ԭ����ʳ��̼ˮ����ռ��������43%��֬��ռ34%���ij�̼ˮ��ռ75%��֬��ռ9%����������ά����3����14�������������ȫ������ͣҩ��ÿ�����ȵ���15��18��λ��5����4��ͣ�á�ԭ���ȵ���40��55��λ��3�˿��Խ��ͼ�����15���º���ã���ʳ��̼ˮ����ռ��������55��60%��ͣҩ��10����7���Բ���ҩ��Ѫ�Dz��ߡ�
Ϊ������������Ч����������ʳ��ά����̼ˮ�����һ��ѧ�߽�8���ȵ��ص������߷�Ϊ���飬̼ˮ���ﶼ��42%����ʳ��ά�ֱ�Ϊÿ��20��3g���ȵ����������䡣10������άѪ��ƽ��1210������ά��ƽ��1690mg��L-1��
����ά���ǵ����������ȵ��ط��ڱȽ�ƽ�ȡ�������ʳ��ά����������һ��������
4.4.2 ȱѪ�����ಡ
��ʳ��ά�Ķ��ٺ�Ѫ�嵨�̴�Ũ����һ����ϵ��Harding��Stare������ʳ����2060����ʳ����2910mg��L-1��
��ʳ��ά���ߣ�������ȡ�ࡣ�ȵ��ط������ӣ���������Ӳ������֭���ڷ�����ų��٣����Ѫ�����̴����ߡ���Щ����ȱѪ�����ಡ�����ء����̽۹���3�ܣ�ÿ��15g��Ѫ�����̴�����13%������֬������44%�����Թ̴�����������17%����֭������33%��
4.4.3 ��ʯ��
��ʬ�췢�ַ�����Һͷ�չ�й��һ���ʯ�İٷ����кܴ���ǰ�߸ߣ����ߵ͡�
��ʯ�γɵ�ԭ���ǵ��̴��ϳɹ��൨֭��ϳɹ��١���ʯ��������ʳ��ÿ�������20��40g��6�ܺ�֭�е��̴��������͡�û�е�ʯ�ĸ�Ů��ÿ�������30g�������º̴��ڵ�֭�е�Ũ��Ҳ���͡�
4.4.4 ���غʹ̼��Գ��ۺ���
��ʳ��ά�ٺ�����١�Ӳ����С�顣�����뷢չ�й��ҵķ����кܴ��������Ӣ��ƽ��ÿ����100��150g���ڸɴ�ũ��470g����X�߹۲�С��ͨ��θ������ʱ�䣬Ӣ����40��100h���ڸɴ�36h��
�̼��Գ��ۺ��ص�֢״������������ʳ�����ˡ�θ���ȡ����ġ����͡������С���ʹ��к������ı��غ�к����ʹ�����᳦��ʹ�ȡ�������Ϊ����ʳ��άȱ������ij����ܸı䡣������ʳ��ά�������õ�Ч������ÿ����������30g��6�ܺ�ʹ�Ĵ����ͳ̶ȡ��ļ��᳦����������١��ر��������е��ڳ�ϰ�ԵĹ��ܣ�ÿ���30g��ԭ��ƽ��3.8����һ�εĿ����̵�2.4�죬ԭ��1.0��һ�εĿ��ӵ�1.72�졣
4.4.5 ���Ҳ�
��ȥ��Ϊ���Ҳ�Ҫ�õ�����ʳ���������෴��70�����߸ijɸ���ά��(����ÿ��3��45g����)��62������36%֢״���ᣬ52%֢״��ʧ����48���������������������ʳ��ά4.5g��Ϊ��6���µ�3�꣬73%�Ļ���֢״�õ����ƣ�33%%��ʹ��ʧ���ð�ο����˫ä������õ����ƽ����
�᳦�������ٺ�ǻ��խ�����γɱպ϶Σ��Ӷ����ӳ��ڵ�ѹ����ͬʱ�����Ӳ��ճ����Ҫ�����ѹ�����ű㣬�����Ҳ�����ʳ��ά���ӷ�����������ˮ���ɽ���Ӳ�Ⱥ�ճ�ȣ���һ���ô���
4.4.6 ��
�����°����ʣ�һ����Ϊ�ڷ��У�������ϸ���Ĵ�л���������ǻ��ĵ�֭�ᡢ�����ĵ����Ρ���������ȡ����ҳ��ڻ����������������
���в�ѧ����˵�����ķ�������ʳ���е����ࡢ֬���������������������أ������Ͷ���ʸ���ء�
������������ż����������������᳦�����ڰ�ϳ���������������������ǣ��ð���������75%����52%�������м������ÿֻ�����������ƽ��6.4���ٵ�2.7������������Ҳ��100%����67%��
��ʳ��ά�ٺ�����٣�����ˮ���٣��°����ʵ�Ũ��������ߣ����ڳ���ͣ��ʱ�䳤��ϸ���������°����ʶ࣬�볦ճĤʱ�䳤������ʳ��ά�������¸��෴��ͬʱ����ά���������Խ��ͣ�����������ʳ�������°��ġ�
4.4.7 ȣ�ݺ����ܲ�
ȣ�ݡ��γ��������������չ�й���ƽ��ÿ�˲���2ö��������10��25��Ӣ��3��4��С��ƽ��3ö��15�������8.4ö��2004����84%�����ܲ���
��������Һ���ڣ��������仺���������������ٸ��������ϵ�ʳ������ڷ�ֹ���ܲ���
4.4.8 �̺��������ż���
����ά��ʳ��ʹ90%�Ҹ߸�ѹ�̴����ߵ�֢״��ʧ����������Լ���п���������ŵ�Ч��������ά��ʳҲ�ɻ�����ѵ�֢״��������������ʳ��ά�Ը��ż���������Ϊ����ѧ�������ܣ�����ά��ʳ�ܷ��������༲���������ۡ�
4.4.9 ��ʳ��ά�����������յ�Ӱ��
��ʳ��ά�ɽ���������ӣ����������Լ������ա���4-15������������Ľ�����Ըơ�þ��Ӱ��Ƚϴ�4-6��5�����Ӻ�4��Ů�ӵĽ������ʳ��ά���߸Ƶĸ�ƽ���൱���ء���������Ӧ���������������6�ܻ������ʱ�䡣�����ʱ���ڣ�ά����D���벻���߲�ɹ̫��������һ����Σ�յġ�
��4-5 ��ά�ض����������յ�Ӱ��
����������
����������
������(g)
������(%)
������(g)
������(%)
��
5.1
99
5.6
98
��
4.6
97
4.3
86
��
13.5
74
13.2
63
��
9.9
81
10.2
74
þ
1.1
73
1.4
59
п
0.12
60
0.13
76
��4-6 ������ͺ�����Ը����յ�Ӱ��
�����*
�����
��������
��ƽ��
��������
��ƽ��
����
580.2��40.0
-26.6��25.2
589.2��24.6
-111.6��26.9
��
425.8��15.3
��
520.3��9.8
-56.8��9.7
���mg��d-1
*�����69�ۣ������ռ����������40��50%��
���⣬��ʳ��ά�ɼ����������ա��������ߣ��ر��������˺��¾��ڵĸ�Ů����ʳ��ά���˹���
4.5 ��ʳ��̼ˮ�������Դ
ʳ����̼ˮ�������Դ������ࣺ����߲ˡ�ˮ�����̺��ǡ������г����ۺ���ʳ��ά�⣬���е����ʡ������ʺ�ά���ء����ࡢ�����ֲ��ĸ��Ϳ龥���ǵ��۵���Դ�������߲˶�����ά�ء������ʡ������ʺ�ά��������֬����ˮ�����������Ǻ����ǡ���ʳ��ά�������ʡ�ά���ء����Ǵ�̼ˮ�����������Ӫ���أ���Ժ���Ӱ��ʳ������������Ӫ���ص���ȡ����
������ʳ����ֻ�������ṩһ��������̼ˮ��������ڳ���ͣ��ʱ�������˫�dz���������ϸ����������ijЩϸ���ܲ���ά����B12������B��ά���ء��˳��������ø����ʧ�������̼�����Ʒ������ijЩ�˵ĸ�к��
һ��ʳ��ɷֱ���ֻ�д���ά�����֡���4-7�г�����ʳ����ʳ��ά������
��4-7 ����ʳ������ʳ��ά�ĺ���
(��λ��ÿ100�˿�ʳ�����еĿ���)
����
����
ľ����
��ʳ��ά����
����
8.0
3.2
44.0
�����
0.7
����
2.7
ȫ�����
1.3
1.2
8.5
ƻ��
0.5
0.5
0
1.4
����
0.4
0.3
0.3
1.4
������
1.0
0.2
����
3.5
���IJ�
0.7
0.7
0.4
2.8
���ܲ�
1.5
0.9
����
3.7
�����
1.MontreuilJ:Glycoproetins,I nComprehensive Biochemistry (Neuberger A&Van DeenenLLM),Vol 19b Parts��,pp.1 ��188,Elsevier,Amsterdam,1982
2.Hakomori SJ:Ann Rev Biochem50:733-64,1981
3.Kuause MV&MahanLK:Carbohydrates,in Food,Nutrition&DietTherapy,7th ed,pp,24 ��29,Saunder,Philadeiphia,1984
4.Silk DBA:Intestinal Absorptionof Nutrients,in Surgical Nutrition(FischerJE),pp.19-25,Little,Brown,Boston,1983
5.Pike RL &BrownMLDigestion,in Nutrition,an Integrated Approach,3rd ed,pp.201-83,John Wiley&Sons,New York,1984
6.Vester JW &RudneyH:Carbohydrates,in Surgical Nutrition (Fischer JE),pp,165 �� 212,Little,Brow Boston,1983
7.Armstrong FB:Pentose phosphatein Biochemistry,2nd,ed,pp,288 �� 93,OxfordUniversity Press, New York,1983
8.Orten Jm&NeuhausOW:Metabolic interrelationship of tisseus,iu Human Biochemistry,10thed,pp,66382,Mosby,St.Louis,1982
9.Levenson SM&SeifterE:Starvation:metabolitc&physiologicres,in Surgical Nutrition(FischerJE),pp.423 �� 445,Little,Brown,Boston,1983
10.Orten JM&Neuhaaus OW:HumanBiochemistry,10th ed.,p.690,Mosby,St.Louis,1982
11.LevyR1:Prevalence&etiology of cardiovascular diseases in Ceicil Textbook ofMedicine(Wyngaarden JB&Smith LH),16th ed.,pp.98 �� 101,Saunders,Philadelphia,1982
12.Albrink MJ&UllrichIH:Effects of dietary fiber on sucrose-induced lipemia in humans,in MetabolicEffects of Utilizable Dietary Carbohydrates(Reser S),pp.221 �� 38,Marcel Dekker,New York,1982
13.Palumbo PJ et al:Short-termsimple carbohydrate loading in patients with coronary artery disease:effectofplasma glucose,serum lipids,plasma levels of in insulin&growthhormone,metabolicrate,&blood pressure,in Metabolic Effects of UtilizableDietary Carbohydrates(Reiser S),pp.315 ��36,MarcelDekker,New York,1298
14.Jeejeehoy KN&MarlissEB:Energy supply in total parenteral nutfition,in SurgicalNutrition(FischreJE),p.647,little,Brown,Boston,1983
15.Williams ER&CahendoMA:Nutrition,Mc Graw-Hill,New York,P.62,1984
16.Pike RL&Brown ML:Theplasma membrane,in Nutrition,an Integrated,Approach,3rd ed,pp.337 ��67,John Wiley&Sons,New York,1984
17.Ashwell &Harford J:Ann RevBiochem51:531��54,1982
18.����������������룺���ķ�������ѧ��195��213ҳ���������������磬������1983
19.Royal Collegeof Physicians:Medical Aspects of Dietary Fibre,pp.1 ��175,Pitman,Kent,1980
4.1.1 ̼ˮ����ķ���
̼ˮ����ֵ��ǡ�˫�ǡ��;��ǡ��������ࡣ�ǵĽ��������֬���ǵ��ס����������ࡣ
(1)���Dz���ˮ ��ɸ����ǵ�̼ˮ����е��ǡ�������3��7��̼ԭ�ӣ���̼ԭ�����Ķ��٣����γ�Ϊ���������졢�ѡ����ǡ����ǺͶ������м��л�����ʽ���ڣ���Ȼ�������������Ǻ����ǡ�
���Ǿ���ȩ����ͪ������ȩ���߳�ȩ�ǣ���ͪ���߳�ͪ�ǡ�
(2)˫��ÿ������ˮ��������ӵ��ǵ�̼ˮ����˫�ǡ�Ӫ�����������˫�������ǡ���ѿ�ǡ��������֡�
(3)�;���ÿ����ˮ���3��8�����ӵ��ǵ�̼ˮ����Ƶ;��ǣ�Ҳ���˰�ˮ���3��10��������20�����ӵ��ǵ�̼ˮ���������һ�ࡣ���ٷ����ࣺˮ������������Ƿ��Ӷ��������ǵij���ѿ�ǵ;��ǣ���3�������Ƿ�����ɵĽ���ѿ���ǣ��ĸ������Ƿ�����ɵĽ���ѿ���ǵȵȡ���һ��ˮ��ʱ������ֹһ�ֵ��ǣ����ӵ;��ǡ�����е��ӵ;���ˮ����������Ǻ�ľ���ǵȣ��˲��������������á��������Լ��ϳɵ��ӵ;��ǣ��к���Ҫ���������á�
(4)����ÿ���Ӹ�����ӵĵ��ǵ�̼ˮ�����Ƕ��ǡ����ٷ֦��Ͼ��ǡ���-�Ͼ��ǡ��Ӷ�������ࡣ��-�ͦ�-���Ѿ��Ƕ��������Ƿ�����ɵģ�������ķ�ʽ��ͬ��ǰ�����Ԧ�-���ռ�����ģ��������æ�-���ռ�����-�Ͼ����е��ۡ���������ԭ���ࡣ����Ӷ���ǵIJ�ֹһ�ֵ��ǣ���ʳ��ά����һ�࣬����ά���⣬��ʳ��ά����������ά�ء����������ࡢľ���ء�
(5)��֬�Ǻ�֬�ʵĽ�������֬���ɷ����ࣺ����֬������֬��
(6)�ǵ��͵����ǹ��彲��̼ˮ����͵����ʽ�Ϻ�Ļ����ﶼ���ǵ��ס��ǵ�������ܶ࣬���й㷺���������á�30����������ü�����ѧ������˲�ͬ�ķ������
1946��Stacey�ѵ����ʺ����ͣ���̼ˮ����Ļ�ѧ��ӦΪ���ij�ճ���ǣ��������ߣ���ѧ��Ӧ�Ե�����Ϊ���ij�ճ���ס�
1962��Gottschalk����̼ˮ���ϵĽṹ�ֳ������ࡣ����֮�����ظ����ֵ�˫�ǵ�λ�ĵ;���������������뵰���ʽ���߳ƶ��ǵ��ף����˰�������Protegolycans,�������һֱ�����������������ǡ���û���ظ�����˫�ǵ�λ�ĵ;��ǵ�λ�ĵ;������뵰�������ϵij��ǵ��ס�
80��������ķ��࣬�ӻ�ѧ�ṹ�������ֵú�ϸ����������ˡ����µ����ۣ���Gottschalk�ķ��෨Ϊ����
ճ���Ǻ�ճ�������֣�����Ӫ��ѧ�������г��֣���������Stacey��������в�ͬ��ճ����(Mucopolyscaccharides)ָGottschalk����ĵ������е�̼ˮ���ﲿ�֣���ư�������(Glycosaminoglycans)��ճ����ָGotrtschalk�����е��ǵ��ס���ʱ����֬���ǵ��͵�����һ��ͳ���ǽ����(Glycoconjugates)��
4.1.2 ̼ˮ����Ļ�ѧ��ɺͽṹ
(1)���ǵ����е�����Ҫ���������ǣ����Ľṹʽ�����ֱ�ʾ��������ͼ4-1��
������������̼ԭ�ӣ����������ּ�����ͼ����ͼ4-1(A)�пɿ�����2��3��4��5��̼ԭ���������ĸ���������ͬ���Dz��Գ�̼ԭ�ӡ�ÿ�����Գ�̼ԭ���������칹�壬�ǻ���������������������ͬ�Ļ����ϰ���ϰ��벮����[ͼ4-1(A)�е�6��̼ԭ��]������̼ԭ�ӵ��ǻ�������ij�Dϵ���ǡ�ͼ4-1(A)�������߿���IJ�����Dϵ�DZر��Ľṹ�����ԣ�ͼ4-1(A)��������D-�����ǡ�
ͼ4-1(A)��ʾ�Ŀ���ʽ�ṹ���Ի�������������5��ԭ�����ǻ������Ƶ���1��̼ԭ�ӵ��ǻ����ϣ�Ȼ�����ɻ�����ͼ4-1(B)����������1��̼ԭ��Ҳ��ɲ��ԳƵĵġ��ǻ�������ijƦ���D-�����ǣ��ǻ�������ijƦ£�D-�����ǡ�
��������̼ԭ���Dz���һ��ֱ���ϵģ�����̼ԭ�ӵĻ��������εġ�Ϊ�����Գ�̼ԭ�ӵĹ��ͣ��ɰ��������ʾ��ͼ4-1(C)������ƽ�洹ֱֽ�棬���߱�ʾ�ڻ�ƽ��ǰ�������ϸ�ߴ��������ڻ�ƽ��ĺ��档ͼ4-1(B)���Ҳ��
ͼ4-1 �����ǵĽṹʽ
�ǻ�д�ڻ���ƽ��֮�£������ǻ�д�ڻ���ƽ��֮�ϡ�������ʾ�Ľṹʽ����HaworthͶӰʽ��
��ʵ�ϣ���������̼ԭ�Ӳ�����һ��ƽ���ϣ�X������Ľ��˵�������������δ��ڵģ���ͼ4-1(D)���������ı�ʾ������д���ѣ�����һ�����HaworthͶӰʽ�����ҳ�����ͼ4-1(E)�������̼���ⶼʡ�ˣ�����һ�����߱�ʾ�ǻ������ڹ�������һ��ͨ�õķ����������Ǽ�������������ǵ���Glc��
��Ȼ����ڵ������ǣ������ڵ��ǡ�˫�ǡ��;��ǻ�����У�����ͼ4-1(D)����ʽ���ڡ�ֻ����ˮ��Һ�У�����Լ1%�Կ�������ʽ���ڡ�
���������ԡ���������У��е���ʹƫ����ƽ���������е�ʹ֮����������ʱ��������(+)��ʾ��������(-)��ʾ���������е�Dϵ���Ƕ��������ġ�����D-�������������ģ�����ȫ���Ǧ���D(+)�����ǡ�һ���ᵽ�������ǣ����ر�ָ�������⣬���Ǧ���D(+)�����ǡ�
������Dϵ�ġ����칹�塢�����ģ�ȫ���Ǧ£�D(-)���ǡ����Ļ������̼ԭ����ɣ���ͬһƽ���ϡ�
ͼ4-2 �£�D(-)����
�������������ǵ��칹�壬����ͬ���ǵ�4��̼ԭ���ϵ��ǻ��ڻ������棬�������ǵ��ڻ������档����Gal��
(2)˫���������ǵĻ�״�ṹ�У���һ��̼ԭ������һ���ǻ������ɺ���һ�����ǻ��Ļ�����ʧˮ����Ϊ���գ��γɵļ������ռ���
������ ��������
��������Ȼ����ڵ�һ��������Щ����Ϊҩ�
��ROH��һ�����Ƿ��ӣ�������ϵľͳ�Ϊ˫�ǡ����������Ƿ��Ӷ��������ǣ��Ԧ�-1��4�Ǹʼ�(����һ�������Ƿ��ӵĵ�1��̼ԭ���ϵ��ǻ��͵ڶ������ѷ��ӵĵ�4��̼ԭ���ϵ��ǻ���ˮ���϶��γɵļ�����д����1��4)��Ϻ�õ�������ѿ�ǡ�
�����Ǧ���D-�����Ǻͦ£�D-�����Ԧ�-1��2���ռ���ϵġ������Ǧ£�D-�������Ԧ�-1��4���ռ��ͦ���D-���������ϵġ�
(3)�;��� ��ѿ���ǵ���ѿ���ǣ����Ǧ���D-�������Ԧ�-1��4�ͦ�-1��6���ռ���ϵġ��ӵ;��ǽ��ṹ�Ƚϸ��ӣ������ǵ����м��Խ��ܡ�
(4)����
�ٵ��۵����������������Ƿ�����ɵġ���ˮ���ɺ�״�����ȵ�60��80��ɷֳ�ֱ�����ۺ�֧�����������֡�ǰ��Լռ15%������Լռ85%��
ֱ������Ҳ�������Ƿ��ӣ��Ԧ�-1��4�ռ�����һ��ֱ������������ͼ4-3��
֧������Ҳ�������Ƿ�����ɣ����Dz����Ԧ�-1��4���ռ�����ֱ�������з�֧����֧�����Ԧ�-1��6���ռ���ϵġ�ÿһ֧��20��30�������Ƿ��ӡ�ͼ4-4������ʾ��ͼ��ͼ4-5��ͼ4-4�п�����ֵķŴ����Ա�ʾ���ѷ��Ӽ�����᷽ʽ��
ͼ4-3 ֱ������ʾ��ͼ
ͼ4-4 ֧������ʾ��ͼ
ͼ4-5 �����Ƿ�����֧�������еĽ�Ϸ�ʽ
����ԭ��ԭ��6000��3000�������Ƿ�����ɣ��ṹʽ��֧���������ƣ�������֧���ࡣÿһ��֧Լ��11��18�������Ƿ��ӡ���֧���Ǧ�1��6���ռ��������Ǧ�1��4���ռ���
�ۺ�������ѿ�;��Ƕ��߶��ǵ���ˮ����м����ṹ���Ц�1��4�ͦ�1��6���ռ���
����ά����ά���Ǧ�-�������Ԧ�1��4���ռ�����ֱ����ÿ��Լ��2800�������Ƿ��ӡ��ṹ��ͼ4-6��ͼ�������Ƿ��ӵĻ�����ǰ���������ֽ��ƽ�洹ֱ�ġ�
ͼ4-6 ��ά�صĽṹʽ
����ʳ��ά
����ά����D-��¶�ǡ�D-ľ�ǡ�D-�����ǡ�D-�������Ԧ�1��4�������������ģ���֧���а������ǡ������Ǻ�������ȩ�ᡣ
��������������ȩ�����ɵģ��������������ǡ��������ǡ������ǡ�ľ�ǡ�
������������ɸ�¶�ǡ�ľ�ǡ�������ȩ�����������ɣ������а����ǡ�
ľ����ľ�ʲ���̼ˮ����DZ�����ľۺ�����ں�������ʳ��άͬʱ���ڣ��������ƵĹ��ã���������Ϊ��ʳ��ά�е�һ�ࡣ
��ʳ��ά�ʹ���ά��ͬ��ֲ����ʳ���У���θ�����������������ʣ�ͳ����ʳ��ά��ʳ����������IJ������Ϊ����ά��������һ����������ʳ��ά����ʧ����õ���ֵ������ʳ��ά������20��50%��
���ǵ�������;��ǺͶ��Dz������ɵ�����ɣ������ɵ��ǵ�����������ɡ���4-1�г������ǵ����ÿ���һ����ʾ�ṹʽ�������ƺͷ��š�
��4-1 ���ǵ�������
���
�ṹʽ
����
����
������
��-D����������
��DGlen
����������
N-��������-��-D����
��DGlcNAc
��ȩ��
��-L-������ȩ��
��ltdu
������
4-����-��-D-������
��DGal-6SO3h
����һ����ɵ;��ǺͶ��ǵĵ��������������Һ�ᣬ���������������ǻ��⣬����������һ�𡣽ṹʽ��ͼ4-7������NeuAc��
�ǵ�ȩ����������������D-������(ͼ4-8)��ȩ������ԭ���ǻ����Ǵ���ɽ�洼(ͼ4-9)��
����֬��֬�Ļ����ṹ���ʰ���(ͼ4-10)�����İ�����֬����(��14��16��̼ԭ��)�������֬���ʰ���������ΪCer�����������ǻ����������������֬���͵;���(�ɼ��������30������ǻ������������)�����������֬����ȷ���ṹʽ������ʮ�֣���������������֬����IJ�ͬ��
ͼ4-7 ��Һ��
ͼ4-8 D-������
ͼ4-9 ɽ�洼
��֬���������͵����ǵ���������һ������ͬ���������ǵ��������ã��в�������֮����
ͼ4-10��֬�Ļ�ѧ�ṹ
���ǵ����е;����ǵ����еĵ��ǻ���������������֣�
���ǣ���������(Ara)��ľ��(Xy1)��
�����ǣ�������(Fuc)��
���ǣ������ǡ������ǡ���¶��(Man)��
�����������ǣ�������������(GlcNAc)����������������(GallNAc)��
��Һ�ᡣ
��ͬ���ǵ��ף�����Щ���ǻ����������е�һ���֣���ɲ�����15������ĵ;��������ٺ͵����ʽ�϶��ɡ��Ǻ���Ϊ1��85%��
�ᵰ�����еľ۰�������һ�����Ƕ��ǡ��������н�����ֵ���ȩ��(����ȩ�����ȩ��)�Ͱ�������(���������ǻ������ǻ������������)��ϰ����˵���н�����ֵ�˫�ǵ�λ���������࣬����4-2��
���������У����������Ȼ���������������������˾۰����Ƕ��Ƕ�۵������ӡ�
�����ж��ٵ��Ƿ��Ӿۺϳɾ۰����ǣ�������Դ����ͬ����������4000��1000000���ϡ�
��4-2 �������еİ����ǵķ�������
�۰�����
˫�ǵ�λ
ȡ����
��������
��ȩ��
��������
������
��DGel
��DGleNAc
Gal;Xyl
4-����������
��DGel
��DGalNAc
GalNAc 6λ���ữ
Gal;Xyl
6-����������
��DGel
��DGalNAc
GalNAc 6λ���ữ
����Ƥ����
��DGal ��LIeU
��DGlcN
����IdU2λ���ữ ����GalNAc4��6λ���ữ��
Gal;Xyl
���������
��DGal ��LIdU
��DGlcN
GlcN���������������ữ�� ����GlcN6λ���ữ�� ����IdU2λ���ữ
����
��DGal `��LIdU
��DGlcN
GlcN���������������ữ�� ����GlcN3��6λ���ữ�� ����IdU2λ���ữ
Gal;Xyl
���������
��DGal
��DGlcNAc
����GleNAc6
GalN;Man;
Fuc;NeuAc
�����ᵥ�����ڣ�һ�㶼�ǵ�����һ�������������������У�������������⣬��ͨ��������-ľ�Ǻ͵����ʵ�˿����������ģ�һ�������Ͽ�������������
4.1.3 ̼ˮ�������������
(1)�ܽ�ȵ��ǡ�˫�ǡ��;��ǡ�����������ˮ�����۲��ܣ���ˮ���Ⱥ����ˮ���ͣ���ɺ�״�����۾��ᴦ�����ɿ��ܵ��ۡ���ԭ�ܷ�ɢ��ˮ�е����ɫ��̬����Һ������ά�ز��ܡ�����������ˮ��������ʳ��ά����ˮ���ͣ���ˮ������Դ����ΧҺ���PH������Ũ�ȵȶ���ͬ�������������5��������������ˮ����ˮ����ʳ��ά�ʺ���״��ϸ����һЩ�����ܴ����ȥ����֬�ǵ�һ����ˮ��֬�ʵ�һ����ˮ���ǵ��͵����ǵ���Ҳ����ˮ��
(2)����������ǵ������100��һЩ�Ǽ��Ǵ�����ȼ���4-3������ת���������ǵ�ˮ�����������Ǻ��Ǹ�50%��
��4-3 �ǵ����
�ǻ���������
���
����
173
ת����
130
����
100
������
74
ɽ�洼
60
��¶��
50
������
32
��ѿ��
32
����
16
4.2 ̼ˮ�����������л
4.2.1 ��������
̼ˮ����Ҫ�������Dz����ա������Ĺ��̾���ˮ��Ĺ��̡���ѿ�ǡ����ǡ����ǡ���ѿ�;��Ƕ������������������Ķ��ǽ�����һ�֣���ԭ���Ƴ�ʳƷʱ�Ѳ������ˡ�
�����ӿ�ǻ��ʼ����ǻ������Һ����ø��ˮ�⽻���1��4�Ǹʼ���������ˮ���1��6���ռ������ڵĦ�1��6���ռ������������Ǻ�������ѿ�;��Ǻ���ѿ�ǡ�
θ��û���������۵�ø����Һ����ø������PH��6.6��6.8����ʳ��û�б�θ���к���ǰ���ܳ�������һ��ʱ�䣬ʹ���ۺ͵;�����������һ���֡�
С��������Һ�Ħ�-����ø�������ú���Һ����ø��ͬ����ֱ��������������ѿ�Ǻ���ѿ���ǣ�֧��������������ѿ�ǡ���ѿ���Ǽ���4��9�������Ƿ�����ɵĶ��Ц�1��6�ռ�����ѿ�;��ǡ�
��ճĤ��Ƥϸ����������ϸ����ÿһϸ��Լ��3000����ë����ë��Ŀռ����Ч�뾶Լ0.4nm��ֻ���������������ܹ�ͨ��������ëĤ�ϵ�ø��Ӧ��Ĥ�ϵ�ø�����֣��٦�1��4����ø���������Ƿ�������������һ��������������������ѿ��ø��ˮ����ѿ�;��ǵĦ�1��6���ռ���������ø���������ǣ��ܦ�-������ø���������ǡ�
�����������������У��ٳ�ǻ�ڵ�������������˫�Ǻ���ѿ�;��ǣ�����ëĤ�ϵ������������ǵ��ǡ�
��ë˫֬�ʲ��(���)������øǶ��˫֬�ʲ��ڣ�����λ����Ĥ�⡣�������������ȫ��Ƕ��Ĥ�ڵ����䵥�ǵĵ����ʣ����������������ղ������̿��Ա����䵰������ϡ�
���䵰���ڽ����������ǰ���Ƚ�ϳ�ǻ�ڵ�Na+���볦ǻ����ǻ��Na+������ϸ���ڣ��ͷŵ������С�Na+���볦ǻ����ǻ�е�Na+Ũ�ȱ�ϸ���ڸߣ��Ե�Ũ���ŵ���Ũ��Ҫ��������������������ATP��Ӧ���ǽ���ϸ����Լ��15%���س�ǻ��25%��ɢ��Ѫ��60%�뿿����Ĥһ�˵���Ĥ�ϵ���һ���嵰��϶��뿪ϸ������һ��ϲ���Na+���������������ǵ��ٶȱ������Ǵӳ�ǻ��������ϸ�����ٶȿ죬���������Dz���������ϸ����������Ӷ����������Ч�ʡ���ʳ�ӵ���ճ��²�ʱ��95%��̼ˮ���ﶼ�������ˡ�
4.2.2 �����
С�����յ�̼ˮ������Ҫ�������ǡ����ǡ����������֣����ž����͵����ࡣ�����ǽ����ϸ���������ᷴӦ����������-6-���ᣬ����ϸ���ڵ�������Ũ�ȿ�ά���ڵ�ˮƽ��ʹ�����Dz��Ͻ����ϸ���������յ��������У�60%�����ڸ��ڴ�л���������ѭ�������ǺͰ������ڸ���ת��Ϊ�����ǡ�
�������ڸ��ھ��ֽ��л�ṩ��������Ҫ������������ĺϳ���ԭ�����ڸ��ڣ����ж����ת���֬�����͵�֬����֯����������
�����ǵķֽ��л�������Σ�
(1)�ͽ������Ƿֽ�ĵ�һ�������ɱ�ͪ�ᡣ��һϵ�з�Ӧ��̼ˮ���������⾭��ĸ���������ɱ�ͪ����ͬ�����Գ�Ϊ�ͽ⡣����ͬ���Ƿ���ʱ��ͪ���һ�����������������ڣ�����ʱ��ͪ�������ɶ�����̼��ˮ������ʱ��ԭ�����ᡣ
�ͽ��ʮ������(ͼ4-11)��Ϊ��Ҫ�������ǵ�6��̼ԭ��һ��Ϊ������ʹ���������ữ�����ܶ��г������ӽ���ȵIJ��֣�Ȼ����ȥ����ɱ�ͪ�ᡣ
������Ӧ����ϸ�����н��С�ÿһ�������ض���ø�μӡ��ڢ١�����������2����ATP��˵�������Ƿֽ��л��ʼʱҪ���������ƶ����ޡ�������������2����ATP�������ѵ���ͪ�ᣬ����2����ATP���ͽ�������������ࡣ
ϸ������NAD+��NADH�����������㶨�������е�NAD+�����NADH���ͽ�͵�ֹͣ��Ҫʹ���Ѽ�������������ͪ�ᣬ�����NADH����ת�Ƶ������������ȥ������ʱ�⾭������������ˮ������ʱ�⽻����ͪ��ʹ������⡣�������ʱ�뿪ϸ����Ѫ����������ȱ���߿ɷ�����л�����ж���
(2)��ͪ����������øa ����ʱ����ͪ����������塣�ڱ�ͪ������øϵ�İ����£��������dz�������������øϵ�İ����£��������dz�������øA��ͬʱ����1molATP��1molNADH��ά����B1B2��������������μӡ���Ӧ�Dz�����ġ�
(3)������ѭ��������øA�ϵ��������������ǵ�̼ԭ�ӣ���������ѭ������2molCO2����������ǵ���������Ӧ���������н��С���һ���̷�9��(ͼ4-12)��
������ѭ���в���3molNADH,1molFADH2��1molGTP��
NADH����ͨ��FAD��ϸ��ɫ�ؽ�������ͬʱ�ų�������ÿmolNADHͨ����һϵ�з�Ӧ���ų���������220kJ(52.6kCal)����Щ������ATP����ʽ����������ÿmolATP�ܲ���30.5Kj(7.3kCal)��������ÿmolNADHRNUD���������������������ϳ�7molATP������ָ�ڱ�״��(25�棬1������ѹ��������Ͳ���Ũ�ȶ���1M/LPH7)��˵�ġ���ģ�����ڵ�����ʱ��ʵ��֤��ֻ�ܺϳ�3molATP�����÷�Ӧʽ��ʾ��
NADH+H++1/2O2
NAD++H2O+����3ADP+3H3PO4+����
3ATP+3H2oǰ��һ����������Ӧ������һ�������ữ������ż����һ�𣬳�Ϊ�������ữ��
ͼ4-11 �ǵĽͽ�
ʵ��Ҳ֤��FADH2����ʱ����2��ATP��GTP������1molATP��������Щ��ֵ����������ȫ����ʱ�������õ������ɹ������£�
�ٽͽ⣺�����ǡ�2��ͪ��+8ATp
�ڱ�ͪ��������������øA��
2��ͪ���2������øA+2CO2+6ATp
��������ѭ����
2������øA��4CO2+24ATp
ͼ4-12������ѭ��
��ÿmol������������������6molCO2ʱ�ɲ���38molATP������������õ�������38��30��1140kJ[(38��7.3)��277kCal]��ÿmol������������6molCO2��ˮʱ�������ܵĸı���2870KJ(686kCal)�����ԣ���еЧ��=1140/2870��10=40%��
�������ñ�״�������ּ���ģ������ڸ�һЩ��Լ60%��
��������ɵ�����Ҳ�ɿ���̼ˮ�����ṩ�����������������������У���������ѭ����ࡣ֬���͵��������Ҳͨ��������ѭ��������ʳ��������ṩ��������90%����������ѭ����
�����Ǵ�л�����һ��;������ͨ�����Ƕ����ᣬ��������һ����ͨ·�����ڲ����˘y���ֽ�������-����ͨ·��
(4)����-����ͨ·��һͨ·Ŀ�����ṩ�ϳɺ�������ĺ��ǣ��Լ��ϳ�֬���ᡢ���̴��������������Ӧ��NADPH(��ԭ��ø��)(ͼ4-13)��������Լ��30%�������Ǿ�����һͨ����л����ϸ������һͨ·Ҳ�ܻ�Ծ������ʧ���ɵ�����Ѫ��ƶѪ����Ӧ����ϸ�����н��С�
����-5-����ȿ����ں��������ϳɣ��ֿ���5-����ľͪ�Ƿ�Ӧ���ɸ���ȩ3-�������-6-���ᡣ��������һ��·��Ҫ��ϸ��������NADPH�ͺ��ǵ���Ҫ������������ϸ��NADPHҪ�ö࣬��ʣ�����ǿɾ�����ȩ-3-�������-6-����ͽ⣬���ɹ���-6-����ת���������-6-�����������һͨ·��
��һͨ·��ά����B1�μӡ�Ҫʹ��·��ͨ����ʳ������������ά����B1��5-�����ͪ��ת��Ϊ������-6-����ķ�Ӧ������������ά����B1�μӡ�Ҫʹ��·��ͨ����ʳ������������ά����B1��Ӫ��״����
(5)��ԭ�����ǵ�����ϸ�����������Ժ���Ŀɴ�������������ϸ������ѹ�����ߣ�ϸ���������Ǻϳɴ���ӵ���ԭ����Ҫ����ʱ��ԭ�ٷֽ������ǡ������ԭ�����������ǣ�����ά��Ѫ�ǵĺ㶨������ԭ�����������ǣ�ֱ�����ڹ���������
ͼ4-13 ����-����ͨ·
4.2.3 ����Ӫ����л�Ĺ�ϵ
��ϰ�������ͣ������Ϳո��������������Ҫ�߱��ܴ���̼ˮ���������ȱ��ʱ��֤������Ӧ�����ױ��졣
�ͺ�Ѫ�����ظߣ��ȵ��ط������ӣ��ȸ�Ѫ���ط��ڼ��١�ʹ����������ǽ�����ࡢ�����֬����֯����ǿø�������������ѵ���������ԭ������ԭ�ĺϳɡ�������ԭ�Ĵ���������ɰ������Ǿ�������DZ�ͪ��ԭ�ɸ���-3-���ᣬ��������øA�ϳɵ�֬���ᣬ����NADPH�ṩ������������֬�����ٺ͵����ʽ�ϳɼ����ܶ�֬������Ѫ�����͵�֬����֯���档�����̼ˮ����Ҳ�ܺϳ�ijЩ�DZ��谱���ᡣ
�ͺ�Լ4h����������ʳ���е������ǡ��ȸ�Ѫ���ط������ӣ��ȵ��ؼ��٣�תΪ���ø���ԭ�Թ���Ѫ�ǡ�����ԭ���������࣬�����ܶ��õIJ���70g,����һ�����ϵ����ġ����ԣ��Ӳͺ�8h��ʼ����������֬�����ṩ���������Ϳɱ���ǣ�����֬������ռ�������ࡣ֬����ֻ�ܹ��ܣ�����ת��Ϊ�����ǡ���ˣ�Ѫ�ǵ���Դ���ò�ת�����ˡ�һ��˵��������������������⣬���������ᶼ���ܳ��ǵģ����������Ĺ۵㿴��ֻ�б�����Ȱ�����������ǵ���Դ����ʱѪ�ǵ�90%���Ա����ᣬ10%���ԹȰ��ᡣ
�����ܰѼ����������ı��������óɱ�ͪ������������ǡ��ո�ʱ�������н���ѪҺ�İ��������ӣ���������30��40%�DZ����ᡣ�����⵰���ʵı����Ậ������10%������ı�������������ģ������Ǿ��ͽ������ͪ�ᣬ�پ�ת�����ñ����ᡣ�����������ǵ�����û�����ӡ����ӵı�����ʵ���������������ᡣ
����ѹȰ���ȥ����æ�ͪ����ᣬ�ٺ������ij��ǰ������������ת�����ö��ñ�ͪ�ᣬ���������ǡ���-ͪ�����Ҳ�ܾ�������ѭ���γ�ƻ���ᣬ���������ǡ�
4.3 ̼ˮ�������������
4.3.1 ��������
(1)������ÿmol��������������ʱ������������2870Kj(686 kcal)�������ǵķ�������180��ÿ�������Dz���2870/180=15.94kJ(686/180��3.81kcal)����ͳ��ȡ������16Kj(4kcal)������������֯���������Ƕ�Ϊ16KJ(3.75kcal)������Ϊ18Kj(4.1kCal)���ҹ�������ҩ��涨������Һ�õ������Ǻ�1mol�ᾧˮ��ÿ�˲���16��180/198��14.5kJ(3��75��180/198��3.41kCal)��������Һʱ�������ṩ������Ӧ��14.5Kj/g(3.41kCal/g)���㡣
(2)��ʳ��̼ˮ������ռ�����İٷֱ�̼ˮ����û�й涨����Ҫ������Ϊ��������������ڶ��ܱ���ǡ���ÿ��ÿ����������ʳ����������̼ˮ����Ϊ50��100g�����������ͪ������֯���ֽ���࣬�Լ������ӣ��ر��������ӣ���ˮ�Ķ�ʧ��
ʳ����̼ˮ����̫��Ҳ���á�����Ӫ��ѧ�ҵ�������������е�55%Ӧ��̼ˮ��������Ӧ�����е���˫���ṩ�IJ�����14%��
�������(��Ҫ�ǵ���)��ͬʱ���ܻ�õ����ʡ�֬�ࡢά���ء������ʡ���ʳ��ά�����뵥˫��(��Ҫ������)ʱ������ȡ�ó������������Ӫ���ء������������ǹ���������ȣ�ݡ���Ѫ�ܼ���������
(3)���ǹ����Ӱ��
�ٹ��IJ������ǹ��࣬��ʳ��ά���٣����IJ��������ʸߡ��籾���������������˵�������Խ��Խ�ͣ������IJ�������ȴ�������ߡ���1900���ÿ10������167.3�ˣ�����1965���237��7�ˡ�
��1909�굽1965�꣬�����˵�ʳ���У�ÿ��ÿ����ȡ�������½�Լ10%�������ʵı仯����֬������������ռ�İٷ�����������9%�������ӵ���Ҫ��������֬����(��������Ϊ����)����ʳ���м������õĵ��̴���������1909��1913��ÿ��ÿ����495mg��1965����518mg������֬���͵��̴��仯���������Եı仯�ǵ���������٣�������˫�dzԵö��ˡ�1909�ꡫ1913��ÿ��ÿ����ȡ̼ˮ����492g����������ռ31.7%��1965��̼ˮ����374g������ռ51.2%��
�ٴ���ʳ��ά�ĽǶȿ���������������������˵�������Ҳ�������١����dzԵ�ˮ�����߲˵��������䣬ˮ����Ʒ�ִ�ƻ����Ϊ��Ը̽ۡ�ƻ���������࣬�̽۵Ĺ��������ڽ�Ƥ�У�һ�㲻�ԡ���һ�ı䵼������ʳ��ά�������ļ��١�
�Ӻ�ķ��棬Yudkin�Ƚ���15�����ҵ���������֬��������֬����ֲ��֬�������͡��������͡��ܵ��ס����ﵰ�ס����ǵ���ȡ�����IJ������ʵĹ�ϵ������ֻ�����dz�����ء�
�ݺ���������ıȽϣ���˵����ȡ�����IJ������ʸߣ���ʳ��άҲ��һ��Ӱ�졣����ʵ���о�Ҳ��֤����һ�㡣
������Cohen�����ƾ���ɫ�кͳ��ڶ�������ɫ�е�Ҳ���˵��������ʺ�̼ˮ�������������˱Ƚ�(��4-4)��˵������������ߣ������ʸߵö࣬��̼ˮ�����������������������г��벻��
��4-4 ������̫��̼ˮ�����������������ʵıȽ�
���
������(kCal)
̼ˮ��������
����
�������ʣ�%
Kcal
�ƾ�10��������
2237
343
1372
6.6
26.4
0.06
�ƾ�25��������
2559
377
1508
63.0
252.0
2.90
���Ƕ�Բ��ã���ô����Ӧ�Զ��٣��Խ����ˣ�ÿ��ÿ��������2g�������ڲ�������Ѫ�ǡ�����������֬���ᡣ���ಡ�������ߣ��������ṩ������ռ��������5%��������
(4)Ӫ������ʱ̼ˮ�����ѡ��ڷ�̼ˮ���ﲹ������ʱ������ѡ����˫�ǡ����蹩��8400Kj(2000kCal)����������ʱ��586g�����˶�ˮ����������3L����˱�ȻҪ�ý�20%����Һ����������ѹ��Ѫ���4�����������͡���ʹ����к����ѡƽ����������������5������ѿ�;��ǣ���25%ʱ����������������������Ը����á����ֵ;��Ǻ������������ա�
��Χ������Һʱ����ѹ���ܸ���500mOsm��kg-1ˮ�����������Ѫ˨�Ծ����ס�10%������(��ѹ500mOsm��kg-1ˮ,)ֻ�ܶ�ʱ��ʹ�á��Ǵ�����ѹ�ϵͣ�������ע���ˮ�洼����20��40%���������ų�������10%ɽ�洼����Χ����Ѫ˨�γɵĿ����ԣ���10%�����Dz�ࡣľ�Ǵ������������Ͳ������������ڵij����������°��ı�����Ҳ��������ѿ�;��ǣ���Ϊ��ͨ�����࣬�;��Dz���ˮ��������ǹ�������֯���á������辭��Χ�����ṩ��������ʱ��ֻ��Ӧ��֬�����������Ʒ��Intralipid����û�ж��ٸ����ã��ѹ㷺�����ٴ���
���ľ�������Һ��ܿ��ܱ�ѪҺ���ͣ����ؿ�����ѹ�����⣬���øߴ�25%��50%������Ũ��Һ��
4.3.2 ����ϸ������֯
ÿ��ϸ������̼ˮ����京��ԼΪ2��10%����Ҫ����֬���ǵ��͵����ǵ���ʽ���ڡ��ֲ���ϸ��Ĥ��ϸ����Ĥ��ϸ�������Լ�ϸ��������С�
ϸ��Ĥ�����㡣����DZ�Ĥ����Ҫ���ڲ��������������ɡ��ڲ���Ĥ��������֬��(��֬����֬)���ж��ɡ�֬�ʵ�֬�����ǷǼ��Եģ��������ڣ��ڲ�����⣬����֬��˫��(ͼ4-14)���ǡ����ἰ�������Ի��Ŷ�����Ĥ�����ࡣ��Щ������Ƕ��֬��˫���ڣ���ϵ�ǵ����ǣ������ἰ�������Ի��Ŷ�����Ĥ�����ࡣ��Щ������Ƕ��֬��˫���ڣ���������Ĥ�ϡ�Ƕ��Ĥ�ڵ��г���άϸ��������ϸ����Ƥϸ���ϵ���������أ�������Ĥ�ϵ��и�ϸ��������������������������ء��������£�֬��˫����Һ��ġ�֬�ʷ��Ӻ͵����Ӷ�������Ĥ���ƶ�����Ĥ����ֵĺ�����ϸ������ͬ����ϸ����Ĥ��������65%��֬��30%��̼ˮ�����5%��
��ÿ��ϸ������̼ˮ�����⣬�ǽ���ﻹ�㷺�����ڸ���֯�С��Ժ�����֯�к�������֬�����ݷֲ��������ϡ������١�θ��Ƣ���Ρ��Ρ����١�����Ĥ����ϸ������ϸ���ȶ�����֬��
�����������������ڵ�ճҺ�����ǵ��ס��Ǻ����е���ճ���ף�Ѫ���е�ǰ���ס���1-����2-����-����-�ס���Ѫøԭ����ά����ԭ���������ף������еļ�״���ء��ټ�״�ټ��ء������ټ��ء��ٺ�ϸ�������أ�ø�еĵ���ø������ø������ø��ˮ��ø�ȶ����ǵ��ס�
������������ڹǡ����ǡ����졢�ʹ�����Ĥ��Ƥ����Ѫ�ܡ�������ؽ�Һ������Һ�С�
�����֯��ϸ������ʣ���Ҫ�ǽ�ԭ�͵���������ɡ�
4.3.3 ������Ϣ
����������һֱ��Ϊ������Ϣ�ǵ����ʺͺ������,̼ˮ������Ҫ�����ǹ������ܺ�����֯����30�����������˴���ʵ��֤�ݣ���1968���ѹ���̼ˮ������Я����Ϣ���Ժ����кܴ�չ����������ڣ�
ͼ4-14 ��Ĥ���ɵ�ʾ��ͼ
��֬��˫�㣬�ڵ�������Ի������ߴ���֬������
bǶ��Ĥ�ڵ���֬������������Ĥ�⡣ cǶ��Ĥ�ڵ��ǵ��ף�����������Ĥ��
d������Ĥ�ϵĵ����ǡ� e���̴� f�����ʷ���
(1)��Ϊ�ǵ���ϸ����ʶ��ı��ϸ����ϸ�����ܹ��ʶ��ġ����ǿ��ϳ��Է��Ǻ��Լ���ͬ�ģ���������һ�����ģ�������������һ����ġ���Ѹ��ࡢ�������̥����ϸ������ͬһ������������ǻ���Լ�����һ����Ϊ�������ϳ�����ڶԷ�ϸ��Ĥ�ϵ������Ǻ��Լ���ͬ�ġ����罫һ���˵�������ֲ����һ��������ȥ�����ܵ������߰�ϸ�����ų���ƻ���ԭ��֮һ���������ϳ�ֲ������ϸ��Ĥ�ϵ��������������IJ�ͬ��
�ڽϼ������У��ɰ��ǵ�ʶ�����ÿ��ø������
��������Щ�ǵ��������һ�������Ҫ���������ϸ���ϻ���ҲҪ�����������ڷ������õ��ǵ���ϸ�������������ڡ����ߵ����������������ϻ������������Ľṹ���˸ı䡣��ϸ����Ĥ�ϵ�������ʶ�����ָı��˽ṹ��������������ǣ�ʹ���ǽ����ϸ���ڣ�����ø����ʹ֮���⡣���ڷ������õĸ�ϸ������ϣ�����������١�
���鶯������࣬������������ʶ���ǵ������壺
������Һ�����ǵ����������˵�������GΪ������;�������ĩ������Һ�ᡣ��Һ�ᱻø���Ժ�ĩ���ǰ����ǡ���ϸ����ʶ���ĩ���ǰ����ǵ�������G���������ĩ������Һ���������G��
��������������Ѫ���ǵ�����֪��21�֣�����ϸ����Ӧ�ĵ����ߴٺ�ϸ�������ء������ݳ��켤�ء���ëĤ�����ټ��ء������أ����嵰����Ѫ��ͭ�����ף�Ѫ���ؽ�ϵ��ס�ά����B12���ݵ��ס�Ƥ������ס�Ѫ���ǵ��ס��������ǵ��ס���Ѫøԭ�ȡ�
������屾��Ҳ���ǵ��ף�������41000������10%��
�ϻ��ĺ�ϸ��Ĥ�ϵ�����Һ�������������ĺ�ϸ��Ϊ�٣��ܱ���ϸ��ʶ��������ɡ������ĺ�ϸ����������ĩ������Һ�ᣬ�����ϣ����������ڷ������á�
�������������ڸ�ϸ��Ĥ�ϣ���������������ס�
�������¶�������ڸ�ϸ��Ĥ�ϣ���������������ס�
�ܸ�¶�ǻ�N-�������������������ڿݷ�ϸ�����档
��Щ���廹������غ�衢Ƣ�ࡢ�κ����С�������ֹ�ĺ�ϸ��Ҳ�ܱ�Ƣ��ʶ�𡢽�Ϻͽ��⡣
̼ˮ����������������ʶ�����ã����������ڱ����ڻ������ȶ������һ���Ӧ�����ٴ����磺A�����ҩ������������Եķ��ӣ������������Ϊĩ�˵ĵ;������ϣ�������ѡ���Եص���ijЩϸ�������ϸ���ϣ�B��ϣ�ȥ����Һ����Ѫ���ǵ��ף��������ر��ⶨ�����������۸�Ӳ�������ס�ԭ���Ըΰ��и�ϸ�������˳̶ȡ����������ĺ�����Һ����������ı�ֵ�����������Ƹ������Ĵ�С��
(2)�п�ԭ������Щ�;����п�ԭ���á��������е�����;�����Ϊ����Ķ��Ľ�Ϻ�ע�������ڣ��ܲ������塣���ֿ�����ʶ����Ϊ��ԭ�ĵ;��ǡ������ṹ���иı䣬����Ͳ�����������DZ������п�ԭ���á�
����Ѫ�ͷ�����Ǻ�ϸ��Ĥ���ǵ�����֬�ĵ;�����ĩ�˵��Ƿ��ӡ�����������14�����Ƿ�����ɣ�ĩ��4����ͼ4-15��A��Ѫ����ĩһ��������N-�������������ǣ�B��Ѫ���ǰ����ǣ�AB��Ѫ�������ֿ�ԭ�����У�O��Ѫ��û�����������Ƿ��ӣ�����ֻ��13�����Ƿ��ӡ�
ͼ4-15��ͬѪ�Ϳ�ԭ�ĵ;�������ĩ���ĸ�����
����ϸ����Ҳ������ԭ���õ���������NILϸ����һ������������ĩ������Һ�ᡣ�����������ϸ�����Ի��Ժ�����ĩ��ȱ���������Һ����ӡ��Ѷ��Ի���NILϸ��������ע����ڷ���������ͬʱ������Ѫ���в�������ȱ��Һ�����֬���������Ͳ����Ϸ�Ӧ�Ŀ��塣���Ŵ���Ĥ�Ϸ�����֬���ǵ����ֶ�Խ��Խ��ϸ���Լ���������ѧ��ҽѧ�Ľ��������ԭ����ķ�Ӧ�����п������ڰ�֢�����ơ�
(3)��ϸ����ϸ����ճ���з�������ϸ����ϸ����ճ����һ��ճ�ŵķ�ʽ�����֣�
��ֱ��ճ��ϸ���������ǻ�ת��ø����֬���ǵ��ף����ø������(�������)��ϣ�ͬʱ���Ǽ�����һϸ������֬���ǵ����ϣ�ʹ������ϸ��ճ����һ����ͼ4-16������ϸ����ø����֬���ǵ��ľ���Զ�������ڶ���һ��ϸ���Ͻ�ϣ�ֻ������һ��ϸ�����ϡ�����ϸ�����߽Ͻ�����������ϣ���������ϸ������ת�ơ�������ϸ����ѪС�塢�����Ͷ��Ի��ij���άϸ����֤ʵ����������ǻ�ת��ø����-��������ø���ƻ�����ճ�����ã�˵��ͼ4-16�е��ǣ������ǰ����ǡ�
��ͨ��ϸ��������е�һ�����ʣ�������ϸ������ѷ���õ�����CSP�ǵ��ף�LETS�ǵ��ס������ǵ��צ�����ճ�����ȡ�����ϸ���ܲ���CSP��LETS�����Ի�����������ϸ��ճ���١����ڶ��Ի��ļ��߳ɹ�ϸ��������ϸ���������м���CSP��LETS�����������ǵ�������
ͼ4-16 ϸ��ճ��ʾ���ͼ
e ����ø G �����ǵ�����֬ S ������
ϸ��ճ��ʧ���ɴ����������˵�ѪС�������һ���ǵ��ף��;�����ĩ������Һ�ᡣѪ�����Ѻ�ѪС���ͨ����Һ��ճ���ڱ�¶������Ѫ����Ƥ�ϣ���ֹѪ���á�BernardSoulier�ۺ�������ѪС��Ĥ�������ǵ������ܵͣ�����ֹѪ��
(4)ϸ���ĽӴ�����ϸ������ʱ������ϸ�����ӵ�һ�����ܶȾͲ������������ֶ��������������ã��нӴ����ơ�����ϸ��ʧȥ�ĽӴ����ƣ�����������ܶȱ�����ϸ���ߵöࡣ�������ƺ�ϸ������������кܴ�Ĺ�ϵ��
ϸ��Ĥ�Ļ�ѧ�ṹ���������ʹ�л�IJ�ͬ��������ͬ����ϸ������ʱ��һЩ�����DZ�¶�ģ�������ϸ�������ڵ���̥ϸ��������ϸ������˿����ʱ��ijЩ����Ҳ�DZ�¶�ģ�������˿���Ѻ�Ķ�ʱ���ڣ��ܺϳ�һ�㵰���ʰ���Щ�����ڸ�������ʹϸ������Ӵ�����Ĥ�ϵ������ỷ��ø�������ӣ�ϸ����cAMPŨ�����ߣ�����Ϣ����ϸ�����ϣ�ֹͣDNA�ĺϳɡ�����ϸ���ܺϳ���㵰���ʣ����ߺϳɱ߱�����øˮ�⣬���Բ�����Ч�ؽӴ����ƶ��������á��ȵ���ø��ˮ������ڸ��õĵ����ʣ�ʹ�����ij���άϸ��ʧȥ�Ӵ����ƶ������������˱��ʰ�����Ƥϸ��������ϸ������ϸ�������ܶ�����cAMP(cAMP��һ���������cAMP������ϸ��)�����ƣ�������һѧ˵��֤�ݡ��������⣬ϸ�����������ṹ�ĸı�Ҳ��ʧȥ�Ӵ����Ƶ�һ��ԭ����֪��������ϸ�����ǵ�����Һ�Ậ�������ӣ�����֬�ĺ������ٵȡ�
4.3.4 ��������
�ǵ��͵������л������á��ؽ�Һ���д��������ᣬ�ǹؽڻ����������Һ�е��ǵ��ף�ʹʳ�������ƶ����ҿɰ���ʳ�Ӻͷ�㣬ʹ��ճĤ���ܻ�е�ͻ�ѧ�IJ��ˡ����������ǵ��ף��з�ֹ֧���ܺͷ�����Ƥ����������������������������������á���ֳϵͳ���ǵ��͵����ǣ��������ھ����˶���������̥�����á�
4.3.5 ���������ʲ�������ø����
��Щ�����ʣ���ø������Һ�е��ǵ��ף�ƽ����������ø�������������ڷ����е������ڱ������ǡ������ṹ�ĸı䣬���M���ٷ��ڵ��ǵ���ȥ��ĩ�˵���Һ������������ĵ���ø������
4.3.6 ����ϸ��Ĥ��ͨ��
�������Ĥ����������������Ի��ţ��ܿ���ˮ���ӡ������Ӻ�С���ӵ��л�����ƶ��ͽ���ϸ���ڲ������跨ʹ�������ܺϳɵ�������ȥ�������ǾͲ��ܽ��뼦�߳���άϸ����ȥ���Ǵ�л�����ϰ���
4.3.7 ���ʽ�ʡ����
ʳ������̼ˮ���ﲻ�㣬���岻�ò��ӵ�����ȡ����������Ϊ������Ҫ�������г�������Ӫ���ء���Ҫ����ȵذѰ��������ڵ����ʺϳɣ�����ȡ���谱�����ͬʱ��һ��Ҫ���㹻��̼ˮ���﹩Ӧ��
4.3.8 ��֤֬���ij������
ʳ����̼ˮ���ﲻ�㣬����Ҫ���ô����֬���������������������֬���������������һ���ȡ�����֬�����࣬��ֽ��л���м����(ͪ��)������ȫ�������������ͪ������ʳ�е�̼ˮ����ɱ�֤����������������
4.3.9 ��ⶾ����
���е�������ȩ���ܽ��һЩ�����Ļ�����Լ�ϸ�������Ķ��صȣ���ͬ�ų����⣬�ⶾ���á�
4.3.1 0��Ϊ�ϳ��������ӵ�ǰ��
������������������̼ˮ�������ϳɵģ������ʡ���ड�ijЩ�����ᡢ߲�������̴��ȡ��������Ǻϳɺ��ᡢ�����ʼ����̴���һЩ�����ʳ���е����������õú��١���ɺ���ĺ��Ǻ��������������������кϳɵġ�
4.3.1 1��ʳ��ά����������
�˲���������ʳ��ά�����᳦��ϸ����ø��ʹ��ά�ء�����ά�غ����ֽ⡣���Դ�����ų�����ά��ֻ��ʳ���е�20��70%������ά��15��45%������Լ10%������Լ70%���ֽ�������⡢������̼�����ᡢ���ᡢ����Ͷ���ȡ��ֽ�ʱҲ���������������������е�һ���֡�����������������ġ�
���䲻��������ʳ��ά������������һ�����������á��ڿ�ǻ����Ӿ����̼���Һ�ķ��ڡ���ʱ���ӳ���Ҳ����θҺ�ķ��ڡ���С��������Ҳ��Ӱ�졣����൱��1004kJ(24kCal)��ƻ����ƻ��֭����һСʱ��δ�ָ�����Ϊƻ��֭�е������պܿ죬�̼����ȵ��صķ��ڣ�ƻ�����й�����ʹ�������գ������˹�����ȵ��ط��ڡ���ʳ��ά�������ӵ�֭�ķ��ڣ�������֭�����ʳ�����ͨ����ʱ�䣬���ӷ�����������ٷ��Ӳ�ȡ�
4.4 ��ʳ��ά��һЩ�����Ĺ�ϵ
�����˲���������������ʳ��ά�����Գ���������Ϊ���dz����ӳ��䶯����ֹ�����⣬�������á�1956��Cleave�����������е�ijЩ��������ʳ���г�ȥ����ʳ��ά(�Ծ�����ۺ�������̼ˮ�������Դ)��������ʮ������ڷ�չ�й��ҹ���������ҽ��������ijЩ�����ڵ��ؾ�����Զ�Ϸ������Ϊ�١�1972��Painter��ʳ�������������������Ҳ��õ����õ�Ч�����������˶���ʳ��ά�����ӡ�
4.4.1 ����ʳ��ά�йصļ����У�
ӡ�ȳ��о����Ծ�����Ϊ����ũ���Dz��ס��ͻ�˹̹�����˳Ծ����棬ũ����ȫ����Ʒ��������������������dz��и���ũ�塣
С���������д���άռ5%ʱ���ָ�Ѫ�ǣ���������>75%ʱ��
�ô���ά��������������в��ٵı��������磬13����ҩ���Ƶ������ˣ�ԭ����ʳ��̼ˮ����ռ��������43%��֬��ռ34%���ij�̼ˮ��ռ75%��֬��ռ9%����������ά����3����14�������������ȫ������ͣҩ��ÿ�����ȵ���15��18��λ��5����4��ͣ�á�ԭ���ȵ���40��55��λ��3�˿��Խ��ͼ�����15���º���ã���ʳ��̼ˮ����ռ��������55��60%��ͣҩ��10����7���Բ���ҩ��Ѫ�Dz��ߡ�
Ϊ������������Ч����������ʳ��ά����̼ˮ�����һ��ѧ�߽�8���ȵ��ص������߷�Ϊ���飬̼ˮ���ﶼ��42%����ʳ��ά�ֱ�Ϊÿ��20��3g���ȵ����������䡣10������άѪ��ƽ��1210������ά��ƽ��1690mg��L-1��
����ά���ǵ����������ȵ��ط��ڱȽ�ƽ�ȡ�������ʳ��ά����������һ��������
4.4.2 ȱѪ�����ಡ
��ʳ��ά�Ķ��ٺ�Ѫ�嵨�̴�Ũ����һ����ϵ��Harding��Stare������ʳ����2060����ʳ����2910mg��L-1��
��ʳ��ά���ߣ�������ȡ�ࡣ�ȵ��ط������ӣ���������Ӳ������֭���ڷ�����ų��٣����Ѫ�����̴����ߡ���Щ����ȱѪ�����ಡ�����ء����̽۹���3�ܣ�ÿ��15g��Ѫ�����̴�����13%������֬������44%�����Թ̴�����������17%����֭������33%��
4.4.3 ��ʯ��
��ʬ�췢�ַ�����Һͷ�չ�й��һ���ʯ�İٷ����кܴ���ǰ�߸ߣ����ߵ͡�
��ʯ�γɵ�ԭ���ǵ��̴��ϳɹ��൨֭��ϳɹ��١���ʯ��������ʳ��ÿ�������20��40g��6�ܺ�֭�е��̴��������͡�û�е�ʯ�ĸ�Ů��ÿ�������30g�������º̴��ڵ�֭�е�Ũ��Ҳ���͡�
4.4.4 ���غʹ̼��Գ��ۺ���
��ʳ��ά�ٺ�����١�Ӳ����С�顣�����뷢չ�й��ҵķ����кܴ��������Ӣ��ƽ��ÿ����100��150g���ڸɴ�ũ��470g����X�߹۲�С��ͨ��θ������ʱ�䣬Ӣ����40��100h���ڸɴ�36h��
�̼��Գ��ۺ��ص�֢״������������ʳ�����ˡ�θ���ȡ����ġ����͡������С���ʹ��к������ı��غ�к����ʹ�����᳦��ʹ�ȡ�������Ϊ����ʳ��άȱ������ij����ܸı䡣������ʳ��ά�������õ�Ч������ÿ����������30g��6�ܺ�ʹ�Ĵ����ͳ̶ȡ��ļ��᳦����������١��ر��������е��ڳ�ϰ�ԵĹ��ܣ�ÿ���30g��ԭ��ƽ��3.8����һ�εĿ����̵�2.4�죬ԭ��1.0��һ�εĿ��ӵ�1.72�졣
4.4.5 ���Ҳ�
��ȥ��Ϊ���Ҳ�Ҫ�õ�����ʳ���������෴��70�����߸ijɸ���ά��(����ÿ��3��45g����)��62������36%֢״���ᣬ52%֢״��ʧ����48���������������������ʳ��ά4.5g��Ϊ��6���µ�3�꣬73%�Ļ���֢״�õ����ƣ�33%%��ʹ��ʧ���ð�ο����˫ä������õ����ƽ����
�᳦�������ٺ�ǻ��խ�����γɱպ϶Σ��Ӷ����ӳ��ڵ�ѹ����ͬʱ�����Ӳ��ճ����Ҫ�����ѹ�����ű㣬�����Ҳ�����ʳ��ά���ӷ�����������ˮ���ɽ���Ӳ�Ⱥ�ճ�ȣ���һ���ô���
4.4.6 ��
�����°����ʣ�һ����Ϊ�ڷ��У�������ϸ���Ĵ�л���������ǻ��ĵ�֭�ᡢ�����ĵ����Ρ���������ȡ����ҳ��ڻ����������������
���в�ѧ����˵�����ķ�������ʳ���е����ࡢ֬���������������������أ������Ͷ���ʸ���ء�
������������ż����������������᳦�����ڰ�ϳ���������������������ǣ��ð���������75%����52%�������м������ÿֻ�����������ƽ��6.4���ٵ�2.7������������Ҳ��100%����67%��
��ʳ��ά�ٺ�����٣�����ˮ���٣��°����ʵ�Ũ��������ߣ����ڳ���ͣ��ʱ�䳤��ϸ���������°����ʶ࣬�볦ճĤʱ�䳤������ʳ��ά�������¸��෴��ͬʱ����ά���������Խ��ͣ�����������ʳ�������°��ġ�
4.4.7 ȣ�ݺ����ܲ�
ȣ�ݡ��γ��������������չ�й���ƽ��ÿ�˲���2ö��������10��25��Ӣ��3��4��С��ƽ��3ö��15�������8.4ö��2004����84%�����ܲ���
��������Һ���ڣ��������仺���������������ٸ��������ϵ�ʳ������ڷ�ֹ���ܲ���
4.4.8 �̺��������ż���
����ά��ʳ��ʹ90%�Ҹ߸�ѹ�̴����ߵ�֢״��ʧ����������Լ���п���������ŵ�Ч��������ά��ʳҲ�ɻ�����ѵ�֢״��������������ʳ��ά�Ը��ż���������Ϊ����ѧ�������ܣ�����ά��ʳ�ܷ��������༲���������ۡ�
4.4.9 ��ʳ��ά�����������յ�Ӱ��
��ʳ��ά�ɽ���������ӣ����������Լ������ա���4-15������������Ľ�����Ըơ�þ��Ӱ��Ƚϴ�4-6��5�����Ӻ�4��Ů�ӵĽ������ʳ��ά���߸Ƶĸ�ƽ���൱���ء���������Ӧ���������������6�ܻ������ʱ�䡣�����ʱ���ڣ�ά����D���벻���߲�ɹ̫��������һ����Σ�յġ�
��4-5 ��ά�ض����������յ�Ӱ��
����������
����������
������(g)
������(%)
������(g)
������(%)
��
5.1
99
5.6
98
��
4.6
97
4.3
86
��
13.5
74
13.2
63
��
9.9
81
10.2
74
þ
1.1
73
1.4
59
п
0.12
60
0.13
76
��4-6 ������ͺ�����Ը����յ�Ӱ��
�����*
�����
��������
��ƽ��
��������
��ƽ��
����
580.2��40.0
-26.6��25.2
589.2��24.6
-111.6��26.9
��
425.8��15.3
��
520.3��9.8
-56.8��9.7
���mg��d-1
*�����69�ۣ������ռ����������40��50%��
���⣬��ʳ��ά�ɼ����������ա��������ߣ��ر��������˺��¾��ڵĸ�Ů����ʳ��ά���˹���
4.5 ��ʳ��̼ˮ�������Դ
ʳ����̼ˮ�������Դ������ࣺ����߲ˡ�ˮ�����̺��ǡ������г����ۺ���ʳ��ά�⣬���е����ʡ������ʺ�ά���ء����ࡢ�����ֲ��ĸ��Ϳ龥���ǵ��۵���Դ�������߲˶�����ά�ء������ʡ������ʺ�ά��������֬����ˮ�����������Ǻ����ǡ���ʳ��ά�������ʡ�ά���ء����Ǵ�̼ˮ�����������Ӫ���أ���Ժ���Ӱ��ʳ������������Ӫ���ص���ȡ����
������ʳ����ֻ�������ṩһ��������̼ˮ��������ڳ���ͣ��ʱ�������˫�dz���������ϸ����������ijЩϸ���ܲ���ά����B12������B��ά���ء��˳��������ø����ʧ�������̼�����Ʒ������ijЩ�˵ĸ�к��
һ��ʳ��ɷֱ���ֻ�д���ά�����֡���4-7�г�����ʳ����ʳ��ά������
��4-7 ����ʳ������ʳ��ά�ĺ���
(��λ��ÿ100�˿�ʳ�����еĿ���)
����
����
ľ����
��ʳ��ά����
����
8.0
3.2
44.0
�����
0.7
����
2.7
ȫ�����
1.3
1.2
8.5
ƻ��
0.5
0.5
0
1.4
����
0.4
0.3
0.3
1.4
������
1.0
0.2
����
3.5
���IJ�
0.7
0.7
0.4
2.8
���ܲ�
1.5
0.9
����
3.7
�����
1.MontreuilJ:Glycoproetins,I nComprehensive Biochemistry (Neuberger A&Van DeenenLLM),Vol 19b Parts��,pp.1 ��188,Elsevier,Amsterdam,1982
2.Hakomori SJ:Ann Rev Biochem50:733-64,1981
3.Kuause MV&MahanLK:Carbohydrates,in Food,Nutrition&DietTherapy,7th ed,pp,24 ��29,Saunder,Philadeiphia,1984
4.Silk DBA:Intestinal Absorptionof Nutrients,in Surgical Nutrition(FischerJE),pp.19-25,Little,Brown,Boston,1983
5.Pike RL &BrownMLDigestion,in Nutrition,an Integrated Approach,3rd ed,pp.201-83,John Wiley&Sons,New York,1984
6.Vester JW &RudneyH:Carbohydrates,in Surgical Nutrition (Fischer JE),pp,165 �� 212,Little,Brow Boston,1983
7.Armstrong FB:Pentose phosphatein Biochemistry,2nd,ed,pp,288 �� 93,OxfordUniversity Press, New York,1983
8.Orten Jm&NeuhausOW:Metabolic interrelationship of tisseus,iu Human Biochemistry,10thed,pp,66382,Mosby,St.Louis,1982
9.Levenson SM&SeifterE:Starvation:metabolitc&physiologicres,in Surgical Nutrition(FischerJE),pp.423 �� 445,Little,Brown,Boston,1983
10.Orten JM&Neuhaaus OW:HumanBiochemistry,10th ed.,p.690,Mosby,St.Louis,1982
11.LevyR1:Prevalence&etiology of cardiovascular diseases in Ceicil Textbook ofMedicine(Wyngaarden JB&Smith LH),16th ed.,pp.98 �� 101,Saunders,Philadelphia,1982
12.Albrink MJ&UllrichIH:Effects of dietary fiber on sucrose-induced lipemia in humans,in MetabolicEffects of Utilizable Dietary Carbohydrates(Reser S),pp.221 �� 38,Marcel Dekker,New York,1982
13.Palumbo PJ et al:Short-termsimple carbohydrate loading in patients with coronary artery disease:effectofplasma glucose,serum lipids,plasma levels of in insulin&growthhormone,metabolicrate,&blood pressure,in Metabolic Effects of UtilizableDietary Carbohydrates(Reiser S),pp.315 ��36,MarcelDekker,New York,1298
14.Jeejeehoy KN&MarlissEB:Energy supply in total parenteral nutfition,in SurgicalNutrition(FischreJE),p.647,little,Brown,Boston,1983
15.Williams ER&CahendoMA:Nutrition,Mc Graw-Hill,New York,P.62,1984
16.Pike RL&Brown ML:Theplasma membrane,in Nutrition,an Integrated,Approach,3rd ed,pp.337 ��67,John Wiley&Sons,New York,1984
17.Ashwell &Harford J:Ann RevBiochem51:531��54,1982
18.����������������룺���ķ�������ѧ��195��213ҳ���������������磬������1983
19.Royal Collegeof Physicians:Medical Aspects of Dietary Fibre,pp.1 ��175,Pitman,Kent,1980